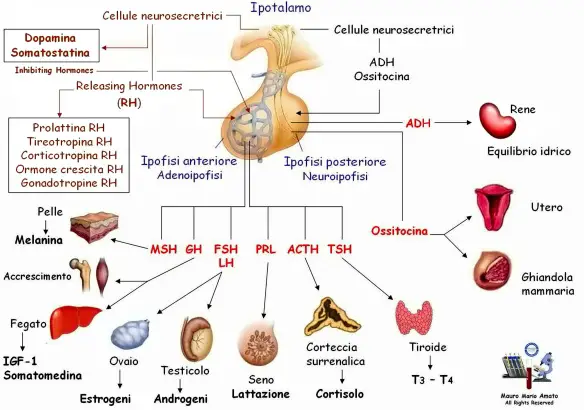
Fonte dell’immagine: dr. Mauro Amato – Centro Analisi Cliniche AMATO sas – Via Tattoli 6/o Molfetta (Bari)
La ghiandola pituitaria (anche chiamata ipofisi) non è più considerata la “ghiandola maestra” come avveniva un tempo: è l’ipotalamo la via finale comune che riceve informazioni praticamente da tutte le altre aree del SNC e che invia informazioni all’ipofisi. L’ipotalamo modula l’attività dei lobi anteriore e posteriore dell’ipofisi in due modi diversi. I neurormoni da esso sintetizzati raggiungono l’ipofisi anteriore (adenoipofisi) direttamente attraverso un sistema vascolare portale specializzato e regolano la sintesi e la secrezione dei sei principali ormoni peptidici dell’ipofisi anteriore. Gli ormoni ipofisari, a loro volta, regolano la funzione delle ghiandole endocrine periferiche (tiroide, surreni e gonadi), oltre all’accrescimento e alla lattazione. Non esistono connessioni neurali dirette tra l’ipotalamo e l’ipofisi anteriore. Al contrario, l’ipofisi posteriore (neuroipofisi) è costituita da assoni che originano dai corpi cellulari di neuroni localizzati nell’ipotalamo. Questi assoni servono come siti di deposito per due ormoni peptidici sintetizzati nell’ipotalamo, i quali a livello periferico hanno la funzione di regolare il bilancio idrico, l’emissione del latte e la contrazione della muscolatura uterina. In alcune specie animali e nel corso dello sviluppo fetale dell’uomo è presente un lobo intermedio localizzato tra il lobo anteriore e quello posteriore, ma nell’uomo adulto, nel quale non è riconoscibile alcuna ghiandola intermedia, le sue cellule sono disperse nel contesto dei lobi anteriore e posteriore. Praticamente tutti gli ormoni prodotti dall’ipotalamo e dall’ipofisi vengono secreti in maniera pulsatile o a poussée, in cui si alternano brevi periodi di inattività e di attività secretoria. Inoltre alcuni ormoni (p. es., l’ormone adrenocorticotropo [AdrenoCorticoTropic Hormone, ACTH], l’ormone della crescita [Growth Hormone, GH] e la prolattina) hanno un preciso ritmo circadiano o giornaliero, con aumento della secrezione durante determinate ore del giorno; altri ormoni (p. es., l’ormone luteinizzante [Luteinizing Hormone, LH] e l’ormone follicolo-stimolante [Follicle-Stimulating Hormone, FSH] durante il ciclo mestruale) hanno ritmi mensili con ritmi circadiani sovrapposti.
Fattori di regolazione
Quando gli ormoni ipotalamici di rilascio e di inibizione raggiungono l’ipofisi anteriore attraverso il sistema vascolare portale, si legano a specifici recettori delle membrane cellulari e danno inizio a sequenze di eventi metabolici che stimolano o inibiscono il rilascio degli ormoni ipofisari nella circolazione generale. Fino a ora sono stati identificati sei neurormoni ipotalamici importanti dal punto di vista fisiologico. Con l’eccezione dell’amina biogena dopamina, sono tutti peptidi di piccole dimensioni. Diversi di essi vengono prodotti a livello periferico, oltre che nell’ipotalamo, ed esplicano la loro funzione anche nel contesto di sistemi paracrini locali, specialmente nel tratto GI. Questi neurormoni possono modulare il rilascio di più di un ormone ipofisario, ma sono specifici nei loro effetti. La regolazione della maggior parte degli ormoni dell’ipofisi anteriore dipende da segnali ipotalamici di tipo stimolatorio positivo; solo la prolattina è sottoposta principalmente a un controllo inibitorio.
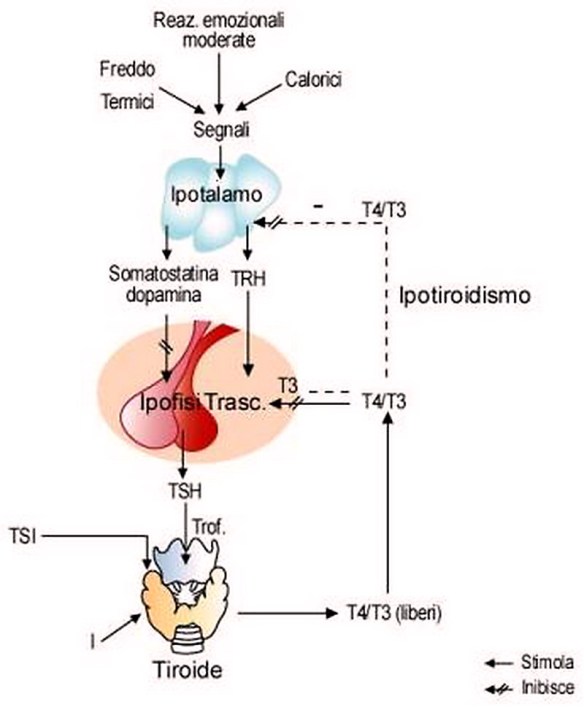
L’ormone di rilascio della tireotropina (Thyrotropin-Releasing Hormone, TRH) induce la sintesi e la secrezione sia dell’ormone tireo-stimolante (Thyroid-Stimulating Hormone, TSH) sia della prolattina. Non è noto se l’induzione del rilascio della prolattina da parte del TRH sia fisiologica. In condizioni patologiche, il TRH può stimolare anche la produzione e il rilascio del GH.
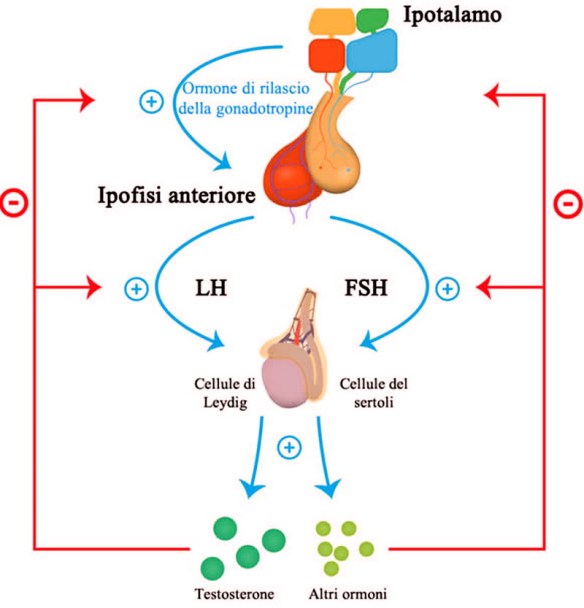
L’ormone di rilascio delle gonadotropine (Gonadotropin-Releasing Hormone, GnRH), conosciuto anche come ormone di rilascio dell’ormone luteinizzante (Luteinizing Hormone-Releasing Hormone, LHRH), stimola la secrezione dell’LH e dell’FSH, sia in condizioni fisiologiche sia quando viene somministrato dall’esterno in maniera pulsatile. Quando viene somministrato GnRH esogeno per infusione continua, il rilascio di FSH e LH viene inizialmente stimolato, ma poco dopo viene inibito a causa dell’inibizione dell’espressione dei recettori ipofisari per il GnRH da parte del GnRH stesso. Questa osservazione ha portato allo sviluppo farmacologico di agonisti del GnRH a lunga durata d’azione, i quali trovano impiego clinico nei casi in cui può essere indicata una castrazione medica. Gli analoghi del GnRH vengono utilizzati con successo per sopprimere la secrezione androgenica negli uomini affetti da carcinoma prostatico, per sopprimere la secrezione steroidea ovarica nelle donne affette da endometriosi e leiomiomi uterini e per sopprimere la secrezione steroidea gonadica nei bambini affetti da pubertà precoce vera. In alcune situazioni il GnRH pulsatile può stimolare anche la secrezione di prolattina.
La somatostatina esercita un controllo inibitorio sulla sintesi e sulla secrezione sia del GH sia del TSH. Il rilascio del GH è stimolato dall’ormone di rilascio dell’ormone della crescita (Growth Hormone-Releasing Hormone, GHRH) e inibito dalla somatostatina; il tasso di produzione del GH dipende dalla forza relativa di questi due stimoli. La somatostatina è inoltre in grado di inibire la secrezione di insulina.
L’ormone di rilascio della corticotropina (Corticotropin-Releasing Hormone, CRH) stimola il rilascio di ACTH dall’ipofisi.
La dopamina è il principale fattore di regolazione della prolattina e ne inibisce la sintesi e il rilascio. Quando il peduncolo ipofisario (che connette l’ipofisi all’ipotalamo) viene sezionato, la secrezione di prolattina aumenta, mentre il rilascio di tutti gli altri ormoni anteroipofisari diminuisce. In alcune circostanze, la dopamina può inibire anche il rilascio di LH, FSH e TSH.
Il peptide intestinale vasoattivo (Vasoactive Intestinal Peptide, VIP) è anch’esso presente nei neuroni dell’ipotalamo e stimola il rilascio della prolattina sia in vivo sia in vitro. Come per il TRH, non è noto se il VIP sia un fattore importante per il rilascio della prolattina in condizioni fisiologiche.
Molte patologie dell’ipotalamo, tra le quali neoplasie, encefaliti e altre lesioni infiammatorie, possono alterare la secrezione dei neurormoni ipotalamici e quindi modificare la funzione ipofisaria. Le sindromi cliniche che conseguono a tali lesioni si presentano come aberrazioni della funzione ormonale ipofisaria. Poiché i vari neurormoni vengono sintetizzati in nuclei diversi all’interno dell’ipotalamo, in alcuni disordini possono essere interessati soltanto uno o pochi neuropeptidi. Nella sindrome di Kallmann, per esempio, un deficit di GnRH ipotalamico causa ipogonadismo. Tuttavia, le lesioni ipotalamiche possono ridurre la secrezione di tutti i neurormoni, determinando un panipopituitarismo secondario con iperprolattinemia e galattorrea (dovuta alla riduzione del rilascio di dopamina). Le lesioni ipotalamiche possono portare anche a ipersecrezione di neurormoni e possono essere responsabili di alcuni casi di pubertà precoce e di sindrome di Cushing.
Funzioni dell’ipofisi anteriore
Le cellule del lobo anteriore (che costituisce l’80% in peso dell’ipofisi e origina da una estroflessione dell’ectoderma della cavità orale) sintetizzano e rilasciano diversi ormoni proteici necessari per l’accrescimento e lo sviluppo normali e stimolano inoltre l’attività di diverse ghiandole bersaglio.
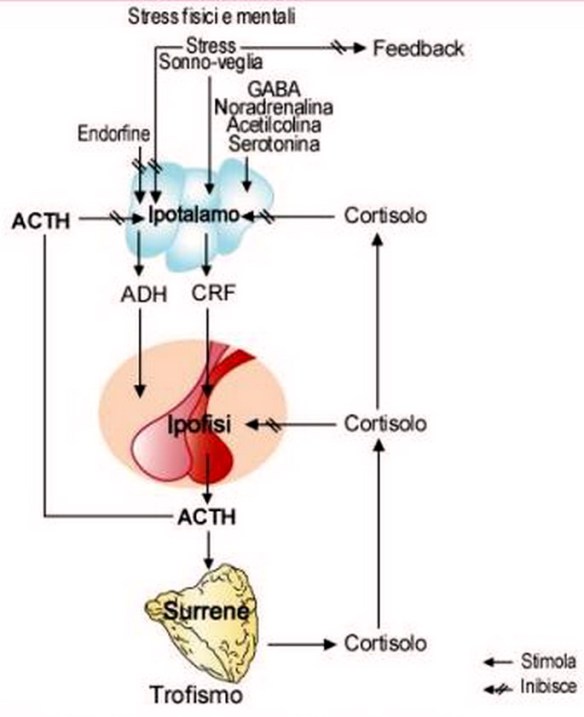
L’ormone adrenocorticotropo (ACTH), conosciuto anche come corticotropina, è un polipeptide a catena singola costituito da 39 aminoacidi. L’attività biologica risiede nei 20 aminoacidi N-terminali. Il CRH è il fattore principale che stimola il rilascio di ACTH e l’ACTH stimola a sua volta la corticale del surrene a secernere cortisolo e diversi androgeni deboli. Il cortisolo e altri corticosteroidi circolanti nel plasma (compresi gli steroidi somministrati a scopo terapeutico) esercitano un feedback negativo sulla secrezione di CRH e ACTH. L’asse CRH-ACTH-cortisolo è fondamentale nella risposta allo stress. In assenza di ACTH, la corteccia surrenale si atrofizza e la secrezione di cortisolo cade praticamente a zero.
Diversi ormoni peptidici vengono sintetizzati a partire da un precursore comune, la pro-opiomelanocortina (POMC), dalla quale originano l’ACTH, la b-lipotropina (b-LPH), l’ormone melanocito-stimolante (Melanocyte-Stimulating Hormone, MSH) a e b, le encefaline e le endorfine. La POMC è presente nel lobo anteriore e nelle cellule derivate dal lobo intermedio dell’ipofisi, oltre che nell’ipotalamo, ma gli ormoni attivi che si formano a partire dalla POMC sono differenti in ciascuna sede in cui essa è presente, in relazione a differenze nell’elaborazione enzimatica. Quindi, l’ACTH e la b-LPH (con una piccola quota di elaborazione aggiuntiva per la formazione di a-LPH e b-endorfina) sono gli ormoni prevalenti sintetizzati nel lobo anteriore. Quasi tutta la b-LPH viene scissa per formare a-LPH e b-endorfina e l’ACTH viene scisso per formare peptide corticotropino-simile del lobo intermedio (corrispondente alla sequenza aminoacidica 18-39 dell’ACTH) e a-MSH (corrispondente alla sequenza aminoacidica 1-13 dell’ACTH) nelle cellule derivate dal lobo intermedio. Inoltre, la sintesi della POMC da parte delle cellule del lobo intermedio sembra essere regolata principalmente dalla dopamina e dalla serotonina, mentre il CRH è l’agente regolatore fondamentale nel lobo anteriore. La POMC e l’MSH possono determinare iperpigmentazione della cute e acquistano importanza clinica esclusivamente nelle patologie in cui i livelli di ACTH sono marcatamente elevati (cioè nel morbo di Addison e nella sindrome di Nelson). Le encefaline e le endorfine sono considerate oppioidi endogeni e si legano ai recettori per gli oppioidi in tutto il SNC, attivandoli.
Gli ormoni glicoproteici ipofisari (cioè l’ormone tireo-stimolante [TSH], l’ormone luteinizzante [LH] e l’ormone follicolo-stimolante [FSH]) e l’ormone placentare gonadotropina corionica umana sono formati ciascuno da una subunità a e una subunità b. Le subunità a di tutti questi ormoni sono identiche, mentre le sequenze delle subunità b sono differenti. Il TSH regola la struttura e la funzione della tiroide e stimola la sintesi e il rilascio degli ormoni tiroidei. La sintesi e la secrezione del TSH sono controllate dall’azione stimolatoria del TRH ipotalamico e dall’azione di feedback negativo svolta dagli ormoni tiroidei circolanti periferici. La sintesi e la secrezione dell’LH e dell’FSH sono stimolate da un unico neurormone ipotalamico, il GnRH (o LHRH), e possono essere soppresse dagli estrogeni. Nella donna, l’LH e l’FSH sono necessari per stimolare lo sviluppo follicolare ovarico e l’ovulazione. Nell’uomo, l’FSH agisce sulle cellule di Sertoli ed è essenziale per la spermatogenesi, mentre l’LH agisce sulle cellule di Leydig del testicolo per stimolare la biosintesi del testosterone.
L’ormone della crescita umano (GH) è un polipeptide a catena singola strutturalmente simile all’ormone placentare somatomammotropina corionica umana, chiamata anche lattogeno placentare umano, e in misura minore alla prolattina. Il GHRH è il principale stimolatore della sintesi e della secrezione del GH e la somatostatina ne è il principale inibitore. Le azioni fondamentali del GH sono la stimolazione dell’accrescimento somatico e la regolazione del metabolismo. L’accrescimento è mediato in larga misura dalla somatomedina C (denominata anche fattore di crescita insulino-simile I [Insulin-like Growth Factor I, IGF-I]), la cui sintesi è controllata dal GH. Nonostante l’IGF-I sia presente in molti tessuti, il fegato è una delle fonti principali dell’ormone. Gli effetti metabolici del GH sono bifasici. Esso esercita inizialmente effetti insulino-simili, aumentando la captazione del glucoso nei muscoli e nel tessuto adiposo, stimolando la captazione degli aminoacidi e la sintesi proteica nel fegato e nei muscoli e inibendo la lipolisi nel tessuto adiposo. Diverse ore dopo la somministrazione del GH questi effetti scompaiono e si manifestano gli effetti metabolici più profondi dell’ormone. Questi effetti più tardivi, che persistono in presenza di un innalzamento prolungato del GH plasmatico, sono anti-insulino simili. La captazione e l’utilizzazione del glucoso vengono inibite, causando un aumento della glicemia e della lipolisi e conseguentemente degli acidi grassi liberi plasmatici. Il GH, i cui livelli aumentano durante il digiuno, è importante nell’adattamento dell’organismo alla mancanza di cibo. Insieme con il cortisolo, l’adrenalina e il glucagone, il GH mantiene il livello di glucoso ematico necessario per l’utilizzazione da parte del SNC e mobilizza i grassi come combustibile metabolico alternativo. I livelli di ormone della crescita diminuiscono con l’età e possono rendere conto in qualche misura della perdita di massa e di forza muscolare che si verifica con l’invecchiamento (sarcopenia).
Le cellule lattotrope secernenti prolattina costituiscono circa il 30% delle cellule dell’ipofisi anteriore. L’ipofisi raddoppia di dimensioni durante la gravidanza, in gran parte a causa dell’iperplasia e dell’ipertrofia di queste cellule. Nel genere umano la funzione principale della prolattina è la regolazione della produzione di latte, ma l’ormone viene rilasciato anche in risposta allo stress e all’attività sessuale. Sebbene la prolattina abbia molti altri effetti in specie diverse, non è chiaro se abbia qualche altra azione fisiologica significativa nell’uomo. La prolattina è l’ormone che più frequentemente viene prodotto in eccesso dai tumori ipofisari.
Funzioni dell’ipofisi posteriore
L’ipofisi posteriore secerne l’ormone antidiuretico (ADH, vasopressina) e l’ossitocina. Entrambi gli ormoni sono peptidi costituiti da nove aminoacidi e vengono sintetizzati in cellule distinte localizzate nel nucleo sopraottico e in quello paraventricolare dell’ipotalamo. Ciascun ormone peptidico viene sintetizzato come parte di un precursore proteico di maggiori dimensioni e rimane legato a una porzione del precursore denominata neurofisina, insieme alla quale viene trasportato lungo gli assoni e depositato in granuli secretori a livello delle terminazioni nervose dell’ipofisi posteriore. L’ADH e l’ossitocina vengono secreti in risposta a impulsi neuronali, si dissociano rapidamente dalle loro neurofisine e vengono prontamente rimossi dalla circolazione, con emivita di circa 10 min. Le neurofisine non esercitano alcun effetto fisiologico conosciuto.
L’ADH e l’ossitocina vengono sintetizzati attraverso la formazione di precursori proteici codificati da geni in singola copia localizzati in posizione contigua sul cromosoma 20. Il proormone per l’ADH, noto come propressofisina o vasopressina neurofisina II, è costituito da quattro porzioni distinte: un peptide di segnale all’estremità amino-terminale, la vasopressina, la sua neurofisina associata e un peptide glicosilato conosciuto come copeptina all’estremità carbossi-terminale. Il proormone per l’ossitocina è simile, ma manca della porzione copeptinica e termina con un singolo residuo di istidina. Con tutta probabilità i proormoni vengono scissi da enzimi proteolitici all’interno dei granuli neurosecretori per dare origine all’ormone attivo e alla rispettiva neurofisina.
L’azione principale dell’ADH è quella di promuovere la ritenzione idrica da parte del rene. Ad alte concentrazioni esso determina anche vasocostrizione. Come l’aldosterone, l’ADH svolge un ruolo importante nel mantenimento dell’omeostasi dei liquidi e del contenuto idrico vascolare e cellulare. Lo stimolo principale per il rilascio di ADH è l’aumento della pressione osmotica dei liquidi corporei, rilevata da osmocettori situati nell’ipotalamo. La deplezione di volume, rilevata da barocettori situati nell’atrio sinistro, nelle vene polmonari, nel seno carotideo e nell’arco aortico, è il secondo maggior stimolo per la secrezione dell’ormone; i segnali di deplezione di volume vengono trasmessi al SNC attraverso il nervo vago e il glossofaringeo. Altri stimoli per il rilascio di ADH sono il dolore, lo stress, l’emesi, l’ipossia, l’esercizio fisico, l’ipoglicemia, gli agonisti colinergici, i b-bloccanti, l’angiotensina e le prostaglandine. Tra gli inibitori della secrezione di ADH vi sono l’alcol, gli a-bloccanti e i glucocorticoidi.
Il diabete insipido è la conseguenza di una carenza di ADH (diabete insipido centrale) o di un’incapacità del rene a rispondere normalmente all’ADH (diabete insipido nefrogenico).
L’ipofisectomia (rimozione della ghiandola pituitaria) generalmente non dà luogo a un diabete insipido permanente, perché un gran numero di neuroni contenenti ADH termina nell’eminenza mediana dell’ipotalamo e continua a funzionare. Per evitare l’insorgenza del diabete insipido è sufficiente la sola presenza di un piccolo numero di neuroni funzionanti contenenti vasopressina.
I due principali organi bersaglio dell’ossitocina sono le cellule mioepiteliali della mammella, che circondano gli alveoli della ghiandola mammaria, e le cellule muscolari lisce dell’utero. In risposta al rilascio di ossitocina stimolato dalla suzione, le cellule mioepiteliali della ghiandola mammaria si contraggono e il latte passa dagli alveoli agli ampi seni lattiferi per essere poi espulso all’esterno (riflesso dell’emissione lattea delle madri che allattano). L’ossitocina stimola la contrazione delle cellule muscolari lisce dell’utero e la sensibilità dell’utero all’ormone aumenta con il procedere della gravidanza, ma le sue concentrazioni plasmatiche non aumentano decisamente durante il parto; il ruolo dell’ossitocina nell’avvio del travaglio non è chiaro. Non è stato identificato alcuno stimolo per la secrezione di ossitocina nel maschio.
Leggi anche:
- Asse ipotalamo-ipofisario: fisiologia e ormoni rilasciati
- Asse ipotalamo-ipofisi-gonade: funzionamento ed ormoni rilasciati
- Asse ipotalamo-ipofisi-testicolo: funzionamento ed ormoni rilasciati
- Asse ipotalamo-ipofisi-surrene: funzionamento ed ormoni rilasciati
- Asse ipotalamo-ipofisi-tiroide: funzionamento ed ormoni rilasciati
- Ipotalamo: anatomia, struttura e funzioni
- Differenze tra ipotalamo, ipofisi, neuroipofisi e adenoipofisi
- Patologie di ipotalamo e ipofisi
- Diabete insipido: cause, diagnosi e trattamento
- Nanismo: sintomi, cura, cause, terapia, diagnosi e prevenzione
- Quando essere troppo alti è una malattia: il gigantismo
- Ipofisi (ghiandola pituitaria): anatomia, funzioni e ormoni secreti
- Ormone della crescita (GH) a che serve e da cosa è prodotto?
- Ormone della crescita (GH): body building e doping in palestra
- Ormone della crescita (GH): effetti avversi nel body building e nello sport
- Sistema nervoso: com’è fatto, a che serve e come funziona
- Sistema nervoso simpatico: funzioni
- Sistema nervoso parasimpatico: funzioni
- Cos’è una ghiandola endocrina? A che servono gli ormoni ed il sistema endocrino?
- Feedback negativo ed omeostasi: spiegazione ed esempi
- Tiroide: dove si trova, com’è fatta e quali funzioni svolge?
- Ipotiroidismo: sintomi, diagnosi, cura farmacologica e consigli dietetici
- Tiroidite di Hashimoto e gravidanza
- Ormoni tiroidei: differenza T3 e T4, valori normali e patologici
- TSH alto, basso e valori normali: qual è il significato clinico?
- Ipertiroidismo: cause, cura, valori diagnosi, sintomi iniziali, conseguenze
- Ipertiroidismo nell’uomo: sintomi, conseguenze sulla libido, cure
- Morbo di Basedow: alimentazione, cura, occhi, si guarisce, rimedi
- Gozzo tiroideo: semplice, tossico, endemico, rimedi, intervento, immagini
- Gozzo tossico nodulare e multinodulare: sintomi, diagnosi e cura
- Alimenti gozzigeni (antitiroidei) vietati nelle alterazioni tiroidee
- Morbo di Plummer (adenoma tossico): sintomi, diagnosi e terapia
- Adenoma tiroideo: sintomi, diagnosi e trattamento
- Eutirox: quando si usa, dosaggio ed effetti collaterali (foglio illustrativo)
- Ipoparatiroidismo e ipocalcemia post chirurgici ed autoimmuni: sintomi e cure
- Tireotossicosi autoimmune, iatrogena, factitia, valori, cura
- Tiroidite acuta, cronica, autoimmune: sintomi e conseguenze
- Tiroidite di Hashimoto: esami, cura, conseguenze, dieta, guarire
- Tiroidite di De Quervain (subacuta): sintomi, dieta, si guarisce, è contagiosa?
- Tiroidite post partum: sintomi, fasi, diagnosi, cura e rischi
- Com’è fatto il cervello, a che serve e come funziona la memoria?
- Cervello maschile e femminile: quali sono le differenze?
- Sistema nervoso autonomo simpatico e parasimpatico: anatomia e funzioni
- Cos’è l’adrenalina ed a cosa serve?
- Adrenalina e “combatti o fuggi”: ecco cosa accade nel nostro corpo quando siamo terrorizzati
- L’altezza media italiana nel 2017 di maschi e femmine
- L’altezza media italiana 2017 da 1 a 19 anni di maschi e femmine
- L’altezza media mondiale nel 2017 di maschi e femmine [TABELLA]
- Altezza: quando si può parlare di nanismo o gigantismo
- Differenza tra midollo osseo e spinale
- Differenza tra sistema nervoso centrale e periferico: anatomia e funzioni in sintesi
- Quanto è alto l’uomo più alto del mondo?
- Quanto è alto l’uomo più basso del mondo?
- Charlotte, la bambina più piccola del mondo
- Tumore benigno e maligno del surrene: sintomi, diagnosi, cura
- Surrene: anatomia, funzioni e patologie in sintesi
- Asse ipotalamo-ipofisi-surrene: funzionamento ed ormoni rilasciati
- Ormone adrenocorticotropo o corticotropina (ACTH): cos’è e quali sono le sue funzioni
- Ormone di rilascio della corticotropina (CRH): produzione e funzioni
- Testosterone basso, alto, valori normali ed interpretazione
- Ormoni estrogeni: cosa sono e quali funzioni svolgono?
- Progesterone: cos’è, a cosa serve, valori e quali funzioni ha in gravidanza?
- Quando la donna ha troppi peli dove non dovrebbero essere: irsutismo, cause, trattamenti e differenze con ipertricosi
- Morbo di Addison: sintomi, immagini, terapia, mortalità, aspettative di vita
- Sindrome di Cushing: immagini, sintomi, diagnosi, cura, guarigione
- Feocromocitoma e surrenectomia: sintomi e conseguenze
- Surrenectomia: tecniche e conseguenze dell’asportazione del surrene
- Iperplasia surrenale congenita: tipi, sintomi, diagnosi, cura
- Iperplasia surrenale acquisita: cause, sintomi, terapia
- Iperandrogenismo femminile: significato, cause, sintomi e terapie
- Iperandrogenismo nell’uomo: significato, cause, sintomi e terapie
- Deficienza di ACTH e cortisolo (ipocortisolismo secondario)
- Deficienza di CRH e cortisolo (ipocortisolismo terziario)
- Malattia di Cushing: cause, sintomi, diagnosi, terapia
- Sindrome di Achard-Thiers
- Sindrome di Waterhouse-Friderichsen: cause e sintomi
Dott. Emilio Alessio Loiacono
Medico Chirurgo
Direttore dello Staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o unisciti al nostro gruppo Facebook o ancora seguici su Twitter, su Instagram, su YouTube, su LinkedIn, su Tumblr e su Pinterest, grazie!
