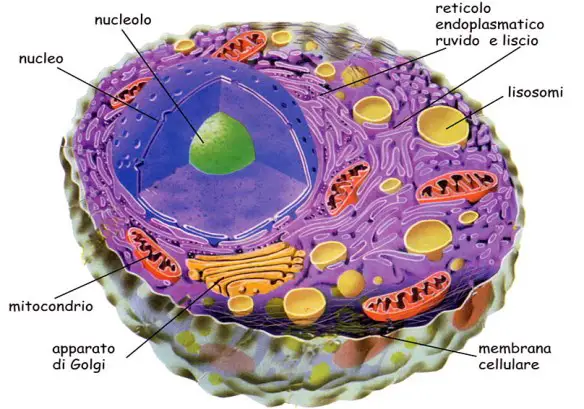
 Grazie al processo di trascrizione (che porta alla formazione di mRNA) e di traduzione (che porta l’mRNA nel ribosoma ad essere usato come “stampo”), all’interno di una cellula vengono sintetizzate numerose proteine. Qual è il destino di tutte queste proteine? Ogni proteina prende una strada diversa in base alla propria funzione. Alcune proteine possono essere secrete (cioè espulse dalla cellula) mentre altre proteine rimangono all’interno della cellula. Queste ultime, in base alla funzione, devono andarsi a localizzare in alcune zone o compartimenti specifici della cellula e tale localizzazione è sotto il controllo genetico. Alcune proteine prodotte dagli eucarioti entrano nel reticolo endoplasmatico, nel mitocondrio, nel cloroplasto o nel nucleo e sono diverse da quelle che invece restano libere nel citoplasma, nel senso che possiedono sequenze specifiche per la destinazione, ovvero sequenze che sono richieste per localizzarle nel comparto cellulare appropriato: le proteine che entrano nel reticolo endoplasmatico possiedono ad esempio sequenze segnale alla loro estremità N-terminale, come vedremo nel prossimo paragrafo.
Grazie al processo di trascrizione (che porta alla formazione di mRNA) e di traduzione (che porta l’mRNA nel ribosoma ad essere usato come “stampo”), all’interno di una cellula vengono sintetizzate numerose proteine. Qual è il destino di tutte queste proteine? Ogni proteina prende una strada diversa in base alla propria funzione. Alcune proteine possono essere secrete (cioè espulse dalla cellula) mentre altre proteine rimangono all’interno della cellula. Queste ultime, in base alla funzione, devono andarsi a localizzare in alcune zone o compartimenti specifici della cellula e tale localizzazione è sotto il controllo genetico. Alcune proteine prodotte dagli eucarioti entrano nel reticolo endoplasmatico, nel mitocondrio, nel cloroplasto o nel nucleo e sono diverse da quelle che invece restano libere nel citoplasma, nel senso che possiedono sequenze specifiche per la destinazione, ovvero sequenze che sono richieste per localizzarle nel comparto cellulare appropriato: le proteine che entrano nel reticolo endoplasmatico possiedono ad esempio sequenze segnale alla loro estremità N-terminale, come vedremo nel prossimo paragrafo.
Le proteine destinate al reticolo endoplasmatico (ER)
Le proteine destinate al reticolo endoplasmatico (ER) hanno specifiche sequenze segnale idrofobiche alle loro estremità amino terminali. Quando una proteina, nel corso della propria sintesi, espone la propria sequenza segnale, una particella per il riconoscimento del segnale (SRP) si lega ad essa e blocca la traduzione ulteriore dell’mRNA. La traduzione resta bloccata finché il complesso polipeptide nascente-SRP-ribosoma-mRNA raggiunge l’ER e vi si lega mediante l’interazione tra la SRP ed una proteina integrale di membrana detta proteina di ancoraggio. La ripresa della traduzione risulta nel trasporto del polipeptide nascente all’interno delle cisterne dell’ER, dove la sequenza segnale viene rimossa. In altre parole, il trasporto di una proteina all’interno dell’ER è un processo cotrasduzionale. Una volta all’interno dell’ER, il complesso di Golgi indirizza poi la proteina alla sua destinazione finale. Anche le proteine destinate al mitocondrio, al cloroplasto oppure al nucleo possiedono sequenze specifiche (sequenze di transito) che le dirigono all’interno dei rispettivi organelli. In questi casi ad essere trasportate all’interno degli organelli sono le proteine ormai completate, quindi in modo posttraduzionale.
Una volta che le proteine mitocondriali o cloroplastiche si trovino all’interno dei rispettivi organelli, la sequenza di transito viene rimossa. Nel caso delle proteine nucleari, tuttavia, la sequenza di localizzazione nucleare resta attaccata, in modo che queste proteine possano venire ancora trasportate nel nucleo dopo la divisione cellulare.
Le proteine trasportate nei mitocondri e nei cloroplasti
Un gran numero di proteine ha come loro destinazione i mitocondri ed i cloroplasti. Tra queste proteine vi sono le proteine ribosomali dei ribosomi degli organelli, alcuni enzimi della catena di trasporto degli elettroni nei mitocondri ed alcuni enzimi per la fotosintesi
nei cloroplasti. Al contrario delle proteine che sono tradotte simultaneamente alla loro entrata nel reticolo endoplasmatico, le proteine destinate ai mitocondri ed ai cloroplasti sono interamente sintetizzate prima che inizi il processo di trasporto, ovvero, i ribosomi che producono queste proteine non si associano con le membrane esterne di nessuno degli organelli. Il tipo di trasporto nel quale la sintesi della proteina viene completata prima della sua importazione nell’organello è detto trasporto posttraduzionale.
Le proteine importate nei mitocondri e nei cloroplasti sono prodotte come precursori che contengono le prima citate sequenze extra alle loro estremità N-terminali dette sequenze di transito, la cui presenza è sufficiente e necessaria per il trasporto. I precursori si associano con specifici recettori sulla membrana esterna dell’organello appropriato e sono di seguito trasportati nell’organello stesso. Una volta all’interno, la sequenza di transito viene rimossa dall’enzima peptidasi per la sequenza di transito.
Le proteine trasportate nel nucleo
Molte proteine hanno come loro destino il nucleo, incluse le proteine ribosomali che sono necessarie per l’assemblaggio dei ribosomi, le proteine cromosomali istoniche e non istoniche ed i molti enzimi richiesti per la replicazione, il riparo, la ricombinazione e la trascrizione del DNA e per il processamento dell’RNA.
Eccetto che per le piccole proteine (<60kDa), il nucleo è estremamente selettivo nei confronti delle proteine a cui permettere l’accesso. Come le altre proteine che abbiamo discusso fino ad ora, anche le proteine nucleari hanno semplici sequenze, chiamate sequenze di localizzazione nucleare, che specificano il loro trasporto all’interno del nucleo. Anche per le proteine destinate al nucleo, come per le proteine mitocondriali e cloroplastiche, il meccanismo di trasporto è di tipo posttraduzionale. Il riconoscimento della sequenza di localizzazione nucleare avviene a livello dei pori della membrana nucleare ed in seguito a tale riconoscimento la proteina viene trasportata nel nucleo attraverso il poro. Al contrario delle altre proteine cornpartimentalizzate che abbiamo discusso, le sequenze di localizzazione nucleare di molte delle proteine nucleari non sono rimosse una volta entrate nel nucleo. La ragione di questo risiede nel fatto che, ad ogni divisione cellulare, l’involucro nucleare viene degradato e si ricostituisce prima della citochinesi, pertanto, dal momento che durante la visione cellulare le proteine nucleari si trovano libere nel citoplasma, devono mantenere la loro capacità di rientrare selettivamente nel nucleo una volta che si sia ricostituito l’involucro nucleare.
Conclusioni
In totale, la sintesi proteica è un complesso processo che richiede l’interazione tra le tre classi maggiori di RNA (mRNA, tRNA e rRNA) ed un gran numero di fattori proteici accessori che agiscono in maniera catalitica nel processo. Con pochissime eccezioni, il codice genetico che specifica l’aminoacido per ogni codone dell’mRNA è lo stesso in ogni organismo, procariota od eucariota. Un gran numero di molecole proteiche possono essere tradotte per traduzione ripetuta di una molecola di mRNA da parte di una sequenza di ribosomi (formanti un polisoma): in questo modo grandi quantità della stessa proteina possono essere prodotte a partire da un singolo gene mediante due passaggi di amplificazione:
- la produzione di mRNA multipli dal gene;
- la sintesi di molte molecole proteiche per traduzione ripetuta di ciascun mRNA.
Per approfondire:
- Sintesi proteica: trascrizione, sintesi dell’RNA, RNA polimerasi I, II e III
- Traduzione dell’mRNA e sintesi delle proteine nei ribosomi
- Differenza tra tra sintesi proteica, trascrizione e traduzione in genetica
Leggi anche:
- Modificazioni post-traduzionali delle proteine: fosforilazione e glicosilazione
- Differenza tra DNA, RNA, ADN, ARN, A-DNA, B-DNA, Z-DNA
- Differenze tra mRNA, tRNA e rRNA
- Differenza tra malattia autosomica dominante e recessiva con esempi
- Differenza tra dominanza semplice, incompleta e codominanza
- Differenza tra allele dominante e recessivo
- Ereditarietà X linked, malattie legate al cromosoma X: significato, esempi
- Differenza tra portatore sano e malato in genetica
- Cosa significa “portatore sano” in genetica e nelle infezioni?
- Differenza tra omozigote ed eterozigote
- Differenza tra genotipo e fenotipo
- Variazione del numero dei cromosomi: aneuploidia, monoploidia, poliploidia
- Aneuplodia, nullisomia, monosomia, trisomia, tetrasomia
- Mutazioni cromosomiche: delezione, duplicazione, inversione, traslocazione
- Delezione in genetica: spiegazione semplice ed esempi
- Duplicazione in genetica: spiegazione semplice ed esempi
- Inversione in genetica: spiegazione semplice ed esempi
- Traslocazione in genetica: spiegazione semplice ed esempi
- Le 10 malattie genetiche più diffuse al mondo
- Sindrome di Down: cause, sintomi in gravidanza e nei neonati
- Sindrome di Turner: cariotipo, cause, sintomi e segni caratteristici
- Sindrome di Rett: cause, sintomi, tipi, diagnosi, stadi, cure, morte
- Sindrome di Klinefelter: cariotipo, cause, sintomi e cura
- Sindrome di Asperger in bambini ed adulti: primi sintomi, terapie
- Sindrome di Noonan: cause, sintomi nel neonato, aspettative di vita
- Sindrome di Bloom: cause, sintomi, diagnosi e terapia
- Sindrome di Pfeiffer: cause, sintomi, diagnosi, cure, prognosi
- Sindrome di Stickler: cause, sintomi, diagnosi, cure, prognosi
- Sindrome di Möbius: cause, sintomi, diagnosi e terapia
- Sindrome di Pierre Robin: cause, sintomi, diagnosi, cure, prognosi
- Sindrome di Wiskott-Aldrich: cause, sintomi, trasmissione, cure
- Sindrome di Lesch-Nyhan: cause, sintomi, trasmissione, diagnosi, cure
- Sindrome di Hughes-Stovin: cause, sintomi, diagnosi e terapia
- Sindrome di Ehlers-Danlos: la malattia dei contorsionisti
- Sindrome di Prader-Willi: cause, trasmissione, sintomi, diagnosi, cure
- Ritardo mentale nei bambini lieve, moderato, grave: si guarisce?
- Sindrome dell’X fragile in uomini e donne: sintomi, aspettativa di vita, cure
- Sindrome dell’idiota sapiente: cause, caratteristiche e sintomi
- Sindrome dell’addome a prugna secca: cause, sintomi, diagnosi, terapie
- Quando una mammella non si sviluppa: la sindrome di Poland
- Diabete insipido: cause, diagnosi e trattamento
- Distrofia muscolare in adulti e bambini: sintomi, cause, diagnosi e cure
- Differenze tra la distrofia muscolare di Duchenne e di Becker
- Emofilia: cos’è, diagnosi, sintomi, tipi, terapia e cura
- Aracnodattilia, segno del pollice e del polso, Sindrome di Marfan e di Beals
- Fibrosi cistica polmonare: cos’è, sintomi in neonati e bambini, cure
- Malattia di Huntington: cos’è, ereditarietà, come si trasmette, età di insorgenza
- Anemia falciforme: cosa significa, cause, sintomi e cure
- Talassemia: cos’è, sintomi, cure, differenti tipi ed alimentazione
- Celiachia: cos’è il glutine, in quali alimenti è contenuto ed in quali no?
- Ectrodattilia: cause, cure ed immagini
- Polidattilia; cause, ereditarietà, sindromica e chirurgia
- Sindrome di Beals e padiglione auricolare “accartocciato”: sintomi e cure
- Differenza tra infiammazione cronica granulomatosa e non granulomatosa
- Malattia granulomatosa cronica: causa, trasmissione, sintomi, cura
- Deficit di glucosio-6-fosfato deidrogenasi e favismo: cause, sintomi, terapie
- Quanti cromosomi hanno esseri umani, scimmie, cani, gatti e topi?
- Quanti cromosomi ha chi è affetto da Sindrome di Down?
- Differenza tra paziente asintomatico e portatore sano
- Differenze tra DNA e RNA: che sono ed a che servono
- Differenza tra DNA ed RNA
- Differenza tra gene e allele
- Cos’è un cromosoma ed a che serve?
- Cos’è un gene ed a che serve?
- Differenza tra malattia genetica, ereditaria e congenita
- Differenza tra genetica ed epigenetica
- Differenza tra genetica e genomica
- Differenza genetica tra uomo e scimmia
- Cosa sono gli alleli ed a che servono?
- Differenza tra cellule eucariote e procariote
- Virus e virioni: cosa sono, come sono fatti, come funzionano e come si riproducono
- Differenza tra cellula aploide e diploide con esempi
- Riproduzione cellulare e ciclo cellulare
- Meiosi: spiegazione di tutte tappe
- Mitosi: spiegazione delle quattro fasi
- Differenza tra mitocondri e cloroplasti
- Differenza tra citosol e citoplasma
- Differenza tra virus e batteri: chi è più pericoloso? Diagnosi, sintomi e terapia
- Organelli (organuli) citoplasmatici della cellula animale: cosa sono ed a che servono?
- Mitocondri: definizione, dimensioni e funzioni
- Citoscheletro: funzioni e struttura
- Ribosomi e reticolo endoplasmatico: cosa sono e che funzioni svolgono?
- Nucleo cellulare: funzioni, dimensioni e membrane nucleari
- Lisosomi: cosa sono? Significato e dimensioni
- Perossisomi: definizione e funzioni
- Membrana plasmatica: definizione e funzioni
- Apparato del Golgi: spiegazione semplice e funzioni
- Citosol: definizione e funzioni
- Differenza tra riproduzione sessuata e asessuata
- Differenza tra vaccini vivi ed attenuati: vantaggi e svantaggi
- Differenza tra immunità specifica ed immunità aspecifica
- Differenza tra linfociti B e T
- Differenza tra immunità umorale e cellulare
- Differenza tra self non self in immunologia
- Differenza tra ciglia e flagelli con esempi
- Differenza tra ciglia e stereociglia: movimento, struttura e funzioni
- Differenza tra ciglia e microvilli: struttura, movimento e funzioni
- Ciclo di Krebs e respirazione cellulare: spiegazione facile e schema
Lo Staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o unisciti al nostro gruppo Facebook o ancora seguici su Twitter, su Instagram o su Pinterest, grazie!
