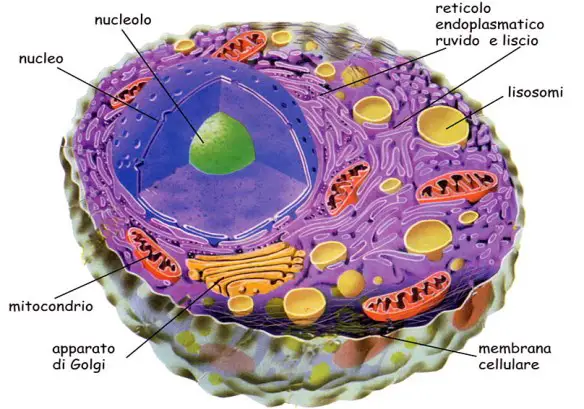
 Il citoplasma non è una massa gelatinosa informe, in cui sono sparsi il nucleo e gli organelli; possiede al contrario una struttura organizzata.
Il citoplasma non è una massa gelatinosa informe, in cui sono sparsi il nucleo e gli organelli; possiede al contrario una struttura organizzata.
Una matrice di proteine fibrose si estende dal nucleo fino alla superficie interna della membrana citoplasmatica, contribuisce a definire la forma della cellula e riveste un ruolo chiave nel movimento e nella divisione cellulare. Questa matrice fibrosa è detta citoscheletro: può anche controllare il movimento degli organelli cellulari e perfino il metabolismo, dirigendo il traffico delle vescicole.
I tre principali componenti del citoscheletro sono: i microtubuli, i filamenti di actina e i filamenti intermedi. I microtubuli e i filamenti di actina sono costituiti da subunità di proteine globulari, che si possono associare e dissociare rapidamente.
Leggi anche:
Il citoscheletro contiene inoltre molte altre specie di proteine accessorie che legano i filamenti fra loro o ad altri componenti cellulari, come la membrana plasmatica, oppure influenzano l’aggregazione delle subunità. Altre proteine accessorie specifiche interagiscono con i filamenti del citoscheletro per produrre movimenti (per esempio, la contrazione muscolare, per opera dei filamenti di actina), o l’azione delle cellule cigliate delle vie respiratorie utilizzando come fonte di energia molecole di ATP.
I microtubuli sono tubicini cavi formati da subunità di una proteina, la tubulina. Nella cellula l’associazione dei microtubuli viene organizzata da strutture specializzate, dette centri di organizzazione dei microtubuli, le quali forniscono una base per l’accrescimento dei microtubuli stessi. Il principale di essi nelle cellule umane, detto centrosoma, si trova vicino al nucleo ed è costituito in genere da una coppia di centrioli, che sono cilindretti cavi la cui parete è formata da 9 triplette di microtubuli. Dai centrioli si organizzano i microtubuli che danno origine al fuso mitotico, che serve da guida per indirizzare i cromosomi ai due poli della cellula in divisione.
La stessa organizzazione dei centrioli la ritroviamo nella cellula in altre strutture: i corpuscoli basali, che hanno il compito di funzionare da impianto per l’organizzazione di ciglia e flagelli. Le ciglia e i flagelli sono sottili appendici filiformi in grado di muoversi, che si presentano sulla superficie di alcune cellule. Essi sono strutturati in un fascio di microtubuli paralleli organizzati in nove coppie poste sulla circonferenza più una coppia al centro, tutto collegato da proteine trasversali. Ciglia e flagelli si differenziano solo per le dimensioni: i flagelli sono più grossi e meno numerosi delle ciglia. La loro funzione primaria è quella di muovere i fluidi sulla superficie di una cellula, o di spingere una cellula isolata attraverso un fluido. Nell’organismo umano, per esempio, vi è un gran numero di ciglia sulla superficie delle cellule epiteliali che tappezzano le vie respiratorie: il loro compito è “spazzare” strati di muco, assieme a particelle di polvere inalate, verso la bocca dove il tutto viene inghiottito ed eliminato. Un tipico esempio di flagello è la coda che permette allo spermatozoo di “nuotare” e spostarsi verso l’uovo per fecondarlo.
Al movimento delle cellule coopera un’altra componente del citoscheletro: i filamenti di actina. Questi sono composti da due subunità diverse e in genere formano una serie di fasci paralleli posti immediatamente al disotto della membrana cellulare. Questi filamenti funzionano da supporti meccanici, in special modo nelle espansioni cellulari. Associati alla miosina, una proteina accessoria, i filamenti di actina formano numerosi sistemi contrattili (per esempio, quelli dei muscoli), che sono responsabili dei movimenti cellulari. I filamenti di actina costituiscono l’anima dei microvilli, piccole espansioni cellulari che formano un orletto a spazzola in molte cellule epiteliali, specie dove è necessaria una grande superficie di scambio come nell’intestino. Fasci di filamenti di actina irrigiditi da legami trasversali contribuiscono nelle cellule capellute dell’orecchio a rivelare il suono. Per poter causare movimento, molto spesso i filamenti di actina sono ancorati alla membrana cellulare tramite una serie di proteine globulari.
Nel muscolo, più filamenti sottili di actina scorrono su un unico filamento più spesso, costituito da miosina, formando strutture complesse che prendono il nome di miofibrille, e nelle cellule muscolari (grosse cellule con più nuclei, risultato della fusione di più cellule) si possono unire più miofibrille a originare un fascio.
La terza componente del citoscheletro è quella dei filamenti intermedi, composta appunto da filamenti che hanno dimensioni intermedie fra quelle dei microtubuli, più grossi, e quelle dei filamenti di actina, più piccoli. I filamenti intermedi sono costituiti da subunità fibrose, associate fianco a fianco per creare una struttura a cordone. Le subunità che formano i filamenti intermedi hanno dimensioni molto diverse, a seconda delle cellule in cui si trovano. La loro funzione è di tipo strutturale, per cui sono particolarmente abbondanti nelle cellule alle quali è necessaria una buona resistenza meccanica.
Leggi anche:
- Differenza tra cellule eucariote e procariote
- Differenza tra cellula aploide e diploide con esempi
- Riproduzione cellulare e ciclo cellulare
- Mitosi: spiegazione delle quattro fasi
- Meiosi: spiegazione di tutte tappe
- Differenza tra disfagia di tipo ostruttivo e di tipo motorio
- Differenza tra disfagia ostruttiva ed occlusione intestinale
- Differenze tra ileo meccanico ed ileo paralitico
- Differenza tra disfagia ai liquidi e ai solidi
- Differenza tra omotrapianto, allotrapianto, xenotrapianto e isotrapianto
- Differenza tra dispnea, apnea e tachipnea
- Differenza tra tachicardia, aritmia, bradicardia e alloritmia
- Differenza tra virus e batteri: chi è più pericoloso? Diagnosi, sintomi e terapia
- Virus e virioni: cosa sono, come sono fatti, come funzionano e come si riproducono
- Differenza tra allele dominante e recessivo
- Differenza tra omozigote ed eterozigote
- Differenza tra gene e allele
- Differenza tra genotipo e fenotipo
- Quanti cromosomi hanno esseri umani, scimmie, cani, gatti e topi?
- Quanti cromosomi ha chi è affetto da Sindrome di Down?
- Cos’è un cromosoma ed a che serve?
- Cos’è un gene ed a che serve?
- Cosa sono gli alleli ed a che servono?
Lo staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o seguici su Twitter, su Instagram o su Pinterest, grazie!
Condividi questo articolo:
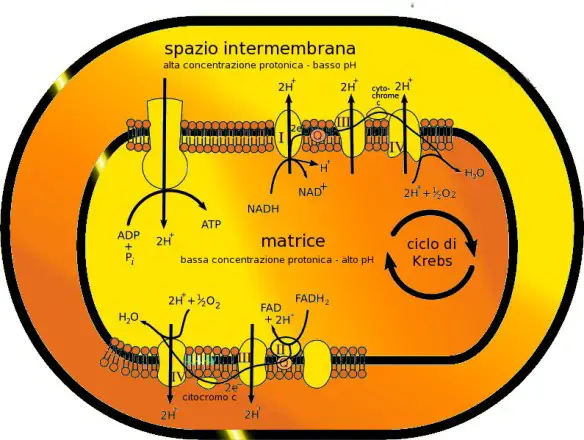 Il mitocondrio (in inglese “mitochondrion”, al prurale “mitochondria”) è una vera e propria “centrale energetica” cellulare: produce l’energia necessaria per molte funzioni cellulari, quali il movimento ed il trasporto di sostanze. I mitocondri contengono gli enzimi necessari per far avvenire le reazioni chimiche che recuperano l’energia contenuta negli alimenti e l’accumulano in speciali molecole di adenosintrifosfato (ATP), nelle quali si conserva concentrata e pronta all’uso.
Il mitocondrio (in inglese “mitochondrion”, al prurale “mitochondria”) è una vera e propria “centrale energetica” cellulare: produce l’energia necessaria per molte funzioni cellulari, quali il movimento ed il trasporto di sostanze. I mitocondri contengono gli enzimi necessari per far avvenire le reazioni chimiche che recuperano l’energia contenuta negli alimenti e l’accumulano in speciali molecole di adenosintrifosfato (ATP), nelle quali si conserva concentrata e pronta all’uso. Le ciglia ed i flagelli
Le ciglia ed i flagelli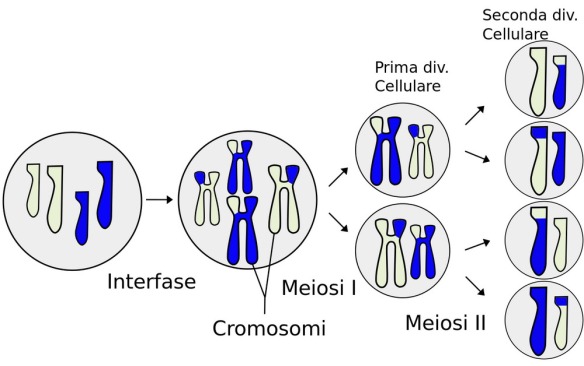 Come abbiamo visto in
Come abbiamo visto in 