 Gli organismi viventi si dividono in organismi unicellulari e pluricellulari.
Gli organismi viventi si dividono in organismi unicellulari e pluricellulari.
- Gli organismi unicellulari possiedono una sola cellula. Esempi sono i virus, i batteri ed i lieviti.
- Gli organismi pluricellulari sono invece formati da più cellule, solitamente specializzate in una data funzione, che collaborano tra loro in modo sinergico. Esempi di organismo pluricellulare sono muffe, funghi ed ovviamente i mammiferi tra cui l’uomo
Leggi anche:
- Differenza tra organismi autotrofi ed eterotrofi
- Differenza tra organismi prototrofi ed auxotrofi
- Differenza tra organico inorganico
- I 12 batteri più pericolosi per l’uomo
- Differenza tra microrganismi, batteri, virus, microbi e germi
- Differenza tra funghi, muffe e lieviti
- Differenza tra capside a simmetria icosaedrica, elicoidale e complessa
- Differenza tra virus e batteri: chi è più pericoloso? Diagnosi, sintomi e terapia
- Virus e virioni: cosa sono, come sono fatti, come funzionano e come si riproducono
- Invasività microbica: la capacità di invadere l’organismo ospite
- Differenza tra invasività clinica e microbica
- Il virus più pericoloso del mondo è più vicino a te di quanto pensi
- Qual è il virus che ha ucciso più persone in assoluto?
- Qual è il virus più letale al mondo?
- Quante persone uccide ogni anno il virus HIV che causa l’AIDS?
- L’insospettabile influenza ogni anno uccide più persone dell’Ebola
- Il virus che uccide mezzo milione di bambini ogni anno
- Febbre gialla, dengue ed altre malattie trasmesse dalle zanzare
- Febbre dengue: sintomi, trasmissione, diagnosi, terapia e prevenzione
- Virus mortali: ecco gli 11 più pericolosi al mondo
- Sierotipo in microbiologia: significato ed importanza per i vaccini
- Differenza tra DNA ed RNA
- Tubercolosi: trasmissione, sintomi, diagnosi e cure in sintesi
- Tubercolosi: cause e patogenesi della malattia
- Mycobacterium tuberculosis: il batterio che causa la tubercolosi
- Sintomi della tubercolosi polmonare ed extrapolmonare
- Come si trasmette la tubercolosi?
- Tubercolosi: diagnosi e progressione della malattia
- Test cutaneo della tubercolina: Test di Mantoux per la tubercolosi
- Trattamento farmacologico per la tubercolosi
- Trattamento della tubercolosi resistente ai farmaci
- Tubercolosi: prognosi, vaccino e strategie di prevenzione
- Differenza tra cellula aploide e diploide con esempi
- Riproduzione cellulare e ciclo cellulare
- Mitosi: spiegazione delle quattro fasi
- Meiosi: spiegazione di tutte tappe
- Differenza tra gene e allele
- Differenza tra allele dominante e recessivo
- Differenza tra omozigote ed eterozigote
- Differenza tra genotipo e fenotipo
- Quanti cromosomi hanno esseri umani, scimmie, cani, gatti e topi?
- Quanti cromosomi ha chi è affetto da Sindrome di Down?
- Cos’è un cromosoma ed a che serve?
- Cos’è un gene ed a che serve?
- Cosa sono gli alleli ed a che servono?
- Differenza tra cellule eucariote e procariote
- Differenza tra coronarografia ed angioplastica
- Differenza tra tumore benigno, maligno, neoplasia, cancro e metastasi
- HIV: sintomi iniziali in donne e uomini
- Differenza tra HIV e AIDS: sono uguali?
- Shock settico e sepsi: sintomi, terapia, conseguenze, si può guarire
- Differenza tra sepsi e Sindrome da risposta infiammatoria sistemica (SIRS)
- Meningite: contagio, sintomi, vaccino, gravità e profilassi
- Endocardite: cause, sintomi, diagnosi e terapie
- Differenza tra batteri Gram negativi e Gram positivi
- Differenza tra batteri bacilli, cocchi, streptococchi e spirilli
- Coprocoltura feci per salmonella: perché e come si fa
- Batteriemia: cura, segni, sintomi, diagnosi ed antibiotici
- I cinque segni cardinali dell’infiammazione
- Differenza tra infezione ed infiammazione: sono la stessa cosa?
- Differenza tra infiammazione cronica granulomatosa e non granulomatosa
- Differenza tra granulomi asettici (da corpo estraneo) e settici
- Si muore di AIDS? Qual è l’aspettativa di vita?
- Sistema immunitario, immunità innata e specifica: riassunto, schema e spiegazione
- Immunodeficienza primaria e secondaria: sintomi, cause e terapie
- Immunità innata (aspecifica): barriere, infiammazione e complemento
- Immunità innata (aspecifica): neutrofili, macrofagi e linfociti natural killer
- Immunità specifica (acquisita): linfociti, T killer, T helper, T γδ, B ed anticorpi
- Immunità specifica (acquisita): memoria passiva, attiva ed immunizzazione
- Immunità specifica (acquisita) umorale e cellulare
- Patologie del sistema immunitario: immunodeficienze, autoimmunità ed ipersensibilità
- Anticorpi: (immunoglobuline): tipi, caratteristiche e funzioni
- Dolore: cos’è, da cosa è causato, quanti tipi di dolore esistono?
- Differenza tra sintomo e segno con esempi
- Differenza tra malattia, sindrome e disturbo con esempi
- Differenza tra organismi aerobi obbligati e facoltativi con esempi
- Differenza tra organismi anaerobi obbligati, facoltativi, microaerofili ed aerotolleranti
- Com’è fatto il cuore, a che serve e come funziona?
- La mammografia: un esame rapido che può salvarti la vita
- Ecografia della tiroide: a cosa serve, come si svolge e come ci si prepara all’esame
- Differenza tra prevenzione primaria, secondaria e terziaria con esempi
- Capire se si ha un tumore: come viene diagnosticato un cancro
- La colonscopia: cos’è, quando si fa, che rischi comporta, come ci si prepara ad affrontarla
- L’ecografia transvaginale esplora gli organi genitali interni femminili
- Mineralometria Ossea Computerizzata (MOC), a cosa serve, come si interpretano i risultati?
- Differenza tra pressione arteriosa e venosa
- Differenza tra atri e ventricoli
- Valvole cardiache: cosa sono, quali sono ed a che
- Differenza tra pressione massima (sistolica), minima (diastolica) e differenziale
- Pressione arteriosa: valori normali e patologici
- Pressione alta (ipertensione arteriosa): sintomi, cause, valori e cure
- Perché la pressione arteriosa alta (ipertensione) è pericolosa?
- Ipertensione: cibi consigliati e da evitare per abbassare la pressione sanguigna
- Qual è la differenza tra arteria e vena?
- Che significa malattia terminale?
- Capire se si ha un tumore: come viene diagnosticato un cancro
- Quanto tempo mi rimane da vivere?
- Fattori di rischio cardiovascolare modificabili e non modificabili
- Cure palliative: cosa sono ed a che servono?
- Eccesso di colesterolo (ipercolesterolemia): perché è pericoloso?;
- Pressione alta (ipertensione arteriosa): sintomi, cause, valori e cure;
- Emoglobina glicata alta, valori normali, IFCC e diabete;
- L’obesità è una malattia;
- Differenza tra malattia acuta e cronica con esempi
- Che significa malattia cronica? Esempi di malattia cronica
- Differenza tra adenocarcinoma e carcinoma con esempi
- Differenza tra cancro e carcinoma con esempi
- Stomie: cosa sono, a che servono, quanti tipi esistono?
- Differenza tra ipertrofia muscolare sarcolplasmatica e miofibrillare
- Ipertrofia muscolare: cosa significa e come si raggiunge
- Cosa sono le metastasi? Tutti i tumori danno metastasi?
- Come nasce un cancro? Cosa sono i cancerogeni e come avviene la cancerogenesi?
- Come prevenire i tumori ed il cancro? I 10 cambiamenti consigliati
- Differenza tra ipertrofia ed iperplasia con esempi
- Differenza tra iperplasia e neoplasia
- Un oggetto che tocchi 150 volte al giorno ha più batteri della tavoletta del wc. Di cosa si tratta e come difendersi
- Differenze tra ileostomia, colostomia e urostomia
- Differenza tra atrofia, distrofia ed aplasia con esempi
- Differenza tra organismi aerobi ed anaerobi con esempi
- Ciclo di Krebs e respirazione cellulare: spiegazione facile e schema
Lo staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o seguici su Twitter, su Instagram o su Pinterest, grazie!
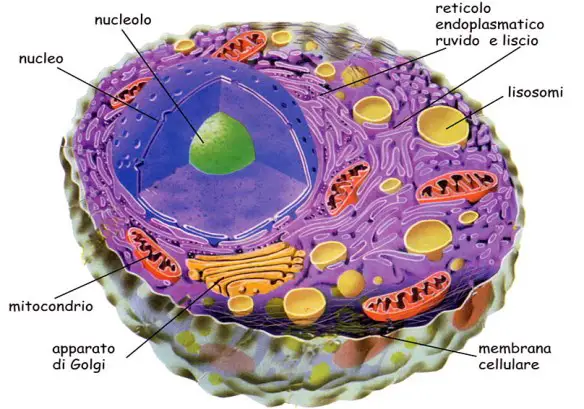 I lisosomi, vescicole che si formano nell’apparato del Golgi, sono essenzialmente sacchetti membranosi (delimitati da una membrana a doppio strato lipidico) che racchiudono enzimi idrolitici, in grado, nella digestione intracellulare, di tagliare in maniera specifica e controllata grosse molecole.
I lisosomi, vescicole che si formano nell’apparato del Golgi, sono essenzialmente sacchetti membranosi (delimitati da una membrana a doppio strato lipidico) che racchiudono enzimi idrolitici, in grado, nella digestione intracellulare, di tagliare in maniera specifica e controllata grosse molecole. Il mitocondrio (in inglese “mitochondrion”, al prurale “mitochondria”) è un organello della cellula eucariote umana. E’ una vera e propria “centrale energetica” cellulare: produce l’energia necessaria per molte funzioni cellulari, quali il movimento ed il trasporto di sostanze. I mitocondri contengono gli enzimi necessari per far avvenire le reazioni chimiche che recuperano l’energia contenuta negli alimenti e l’accumulano in speciali molecole di adenosintrifosfato (ATP), nelle quali si conserva concentrata e pronta all’uso. Il compito dei mitocondri è quello di completare la demolizione delle molecole ingerite come fonte di energia. Infatti, nel citosol gli zuccheri vengono demoliti con reazioni che non utilizzano ossigeno, per cui la digestione è parziale e la resa in energia bassa. Nei mitocondri il metabolismo degli zuccheri (ma anche quello dei lipidi) si completa con la loro ossidazione (ciclo di Krebs). I prodotti di questa reazione vengono utilizzati dalla catena di trasporto degli elettroni per produrre molecole ad alta energia (ATP). In questo modo, l’energia immagazzinata nelle molecole di ATP è molto più alta: infatti da ogni molecola di glucosio vengono prodotte 36 molecole di ATP, mentre la glicolisi a livello del citosol ne produce soltanto 2.
Il mitocondrio (in inglese “mitochondrion”, al prurale “mitochondria”) è un organello della cellula eucariote umana. E’ una vera e propria “centrale energetica” cellulare: produce l’energia necessaria per molte funzioni cellulari, quali il movimento ed il trasporto di sostanze. I mitocondri contengono gli enzimi necessari per far avvenire le reazioni chimiche che recuperano l’energia contenuta negli alimenti e l’accumulano in speciali molecole di adenosintrifosfato (ATP), nelle quali si conserva concentrata e pronta all’uso. Il compito dei mitocondri è quello di completare la demolizione delle molecole ingerite come fonte di energia. Infatti, nel citosol gli zuccheri vengono demoliti con reazioni che non utilizzano ossigeno, per cui la digestione è parziale e la resa in energia bassa. Nei mitocondri il metabolismo degli zuccheri (ma anche quello dei lipidi) si completa con la loro ossidazione (ciclo di Krebs). I prodotti di questa reazione vengono utilizzati dalla catena di trasporto degli elettroni per produrre molecole ad alta energia (ATP). In questo modo, l’energia immagazzinata nelle molecole di ATP è molto più alta: infatti da ogni molecola di glucosio vengono prodotte 36 molecole di ATP, mentre la glicolisi a livello del citosol ne produce soltanto 2.