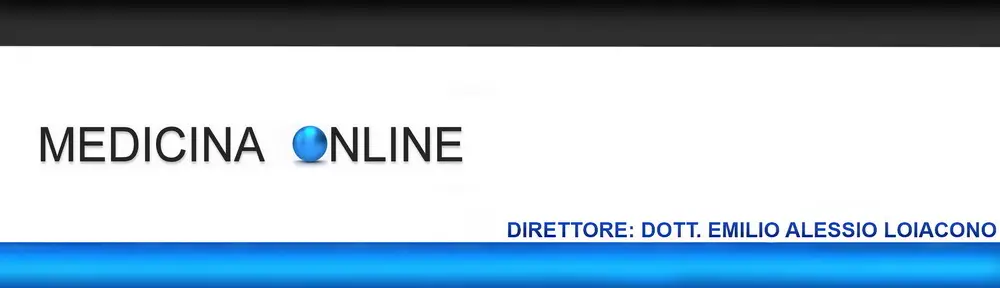
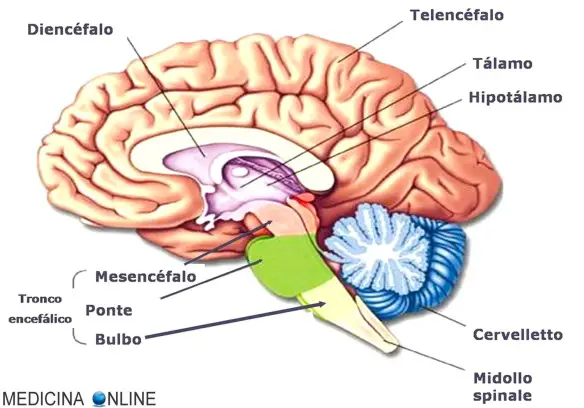
 I meccanismi encefalici che regolano il ritmo circadiano, i cicli e le fasi del sonno e con essi le modificazioni dello stato di coscienza, del comportamento e delle condizioni umorali, endocrine e vegetative sono stati uno dei temi più appassionanti della neurofisiologica degli ultimi 70 anni. Ciò nonostante non è stata trovata ancora una risposta sicura per uno dei più importanti dilemmi, precisamente se il sonno debba considerarsi fenomeno «passivo», consegua cioè alla caduta di un «tono» che mantiene la veglia, ovvero sia fenomeno «attivo», consegua cioè all’attività di uno o più centri cerebrali (Steriade e coll.).
I meccanismi encefalici che regolano il ritmo circadiano, i cicli e le fasi del sonno e con essi le modificazioni dello stato di coscienza, del comportamento e delle condizioni umorali, endocrine e vegetative sono stati uno dei temi più appassionanti della neurofisiologica degli ultimi 70 anni. Ciò nonostante non è stata trovata ancora una risposta sicura per uno dei più importanti dilemmi, precisamente se il sonno debba considerarsi fenomeno «passivo», consegua cioè alla caduta di un «tono» che mantiene la veglia, ovvero sia fenomeno «attivo», consegua cioè all’attività di uno o più centri cerebrali (Steriade e coll.).
Le ormai classiche osservazioni cliniche di Von Economo e quelle sperimentali di Hess indicavano il sonno come fenomeno attivo dipendente dal diencefalo, mentre il preparato sperimentale cerveaux isolée di Bremer induceva a considerarlo fenomeno passivo conseguente a deafferentazione corticale. Fenomeno passivo è stato considerato anche da Moruzzi dopo la scoperta dell’ARAS (ascending reticular activating system), Successivamente però nella stessa scuola del Moruzzi è stato dimostrato per mezzo dei modelli sperimentali «medio-pontino pretrigeminale» e «pretrigeminale emipontino» che nell’ARAS sono comprese strutture sia a funzione ipnogenica, sia atte a mantenere lo stato di veglia; in questo modo si è rivalutata la teoria del sonno come fenomeno «attivo», Jouvet ha confermato il ruolo attivo di certe porzioni dell’ARAS nell’indurre i due tipi di sonno individuando peraltro alcune strutture specifiche, precisamente illocus coeruleus e il rafe con funzione primaria nell’instaurazione del sonno. A questo Autore va il merito di avere iniziato l’era neurochimica nello studio del sonno, stabilendo che l’effetto delle due strutture citate, quanto a sonno REM, si realizza mediante neurotrasmettitori specifici, l’acetilcolina e la norepinefrina.
Nel 1991 Jones, rivisitando in modo aggiornato le ricerche sperimentali, lesionali, elettrofisiologiche e farmacologiche, nonché le osservazioni cliniche relative al substrato strutturale del sonno REM, ha trovato consistenti prove sul ruolo critico che i neuroni colinergici del tegmento dorso-laterale ponto-mesencefalico esercitano nell’induzione del sonno REM e l’apparente modulazione che su di essi svolgono i neuroni noradrenergici del locus coeruleus e quelli serotoninergici del rafe.
D’altra parte Denoyer e coll. hanno dimostrato che lesioni neurotossiche (distruzione cellulare con risparmio delle vie assonali) dell’ARAS mesencefalica e dell’ipotalamo posteriore determinano una perdita di coscienza solo per pochi giorni (a differenza del coma provocato da lesioni elettrolitiche della stessa regione), ciò suggerendo che esistono sistemi di proiezione paralleli e compensatori di quelli neuronali dell’ARAS, capaci di ristabilire e mantenere la veglia. Questi sistemi con effetto attivante corticale ritmico e oscillatorio sono passati in rassegna da Steriade e coll. Comunque altrettanto recentemente l’interesse si è spostato dal tronco encefalico al diencefalo, struttura che gli esperimenti di Hess avevano indicato come sede del centro del sonno. Il talamo può essere chiamato in causa per la caratteristica attività oscillatoria e ciclica dei suoi neuroni, trasmessa alla corteccia cerebrale e imposta ad essi dal nucleo reticolare con neuroni GABAergici, attività cui è attribuito lo spindling ricorrente nella fase II del sonno umano e con esso la selezione, la modulazione e il trasferimento delle afferenze sensitivo-sensoriali, dunque gli stessi differenti stati funzionali di veglia e di sonno NREM.
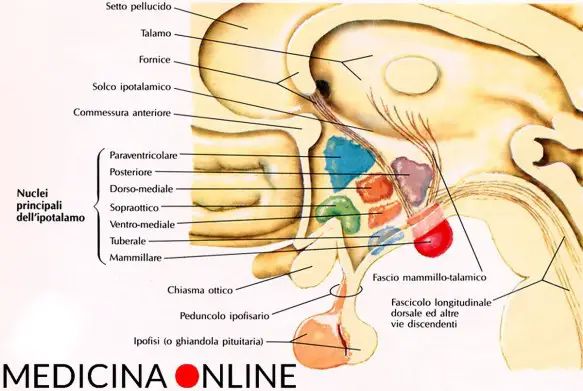
Quanto all’ipotalamo, esso non solo è stato di recente rivalutato nel suo ruolo induttore e regolatore del ritmo del sonno, ma è stato anche proposto come struttura di integrazione reciproca tra sonno NREM e termoregolazione. I neuroni termosensitivi dell’area preottica ipotalamica anteriore sarebbero ad un tempo responsabili dell’omeostasi corporea e dell ‘induzione del sonno NREM. A sua volta il nucleo ipotalamico sovraottico risulta essere l’orologio biologico che governa il ciclo circadiano sonno-veglia. Nel ratto la lesione di questo nucleo determina infatti una sequenza irregolare e aritmica dell’alternanza sonno-veglia e rende inefficace l’effetto ipnotico del triazolam. Con tutta probabilità lo stesso nucleo ha un effetto trigger sul sonno REM.
Va infine segnalato che negli anni recenti è stato documentato l’effetto positivo della melatonina sull’induzione del sonno e sul senso di riposo che ad esso consegue. Come è noto, la melatonina è l’ormone della ghiandola pineale, o epìfisi, sino a 50 anni fa considerata organo vestigiale, attualmente rivalutata nella sua funzione endocrina ad effetto ubiquitario, ma di notevole interesse nell’alternanza sonno-veglia per il fotoperiodismo dei pinealociti nei ritmi circadiani.
Altre recenti osservazioni che sconfinano nella neurochimica del sonno, ma che vengono qui citate perché indicative per le conclusioni che seguono, riguardano i sottotipi di recettori muscarinici, in particolare quelli M2 che risultano interessati nel sonno REM, il ruolo delle prostaglandine cerebrali nel controllo della veglia e del sonno e infine l’effetto dei peptidi muramilici, prodotti dallo Staphylococcus aureus e dai gonococchi, capaci di indurre sonno NREM e innalzamento della temperatura, stimolando la produzione di interleuchina 1 (IL-1).
Concludendo, si può osservare come la distribuzione lungo l’asse troncoencefalico-diencefalico di centri e di sistemi odologici ad effetto ipnogenico, regolatore del ritmo e del mantenimento del sonno, assegna alla corteccia cerebrale un ruolo passivo nel processo ipnico. In realtà, pur non essendo necessaria al realizzarsi dei quadri morfeici, come è stato dimostrato sperimentalmente in animali decorticati, essa ugualmente esercita un’ azione di controllo sui centri sottocorticali ipnoinducenti e ipnoregolatori come si può facilmente intuire considerando il parziale controllo che volontariamente si può esercitare sul sonno e il fatto che la stessa attività mentale conativa lo può ritardare o abolire temporaneamente.
Inteso sia come fenomeno unitario di alternanza alla veglia, sia nelle sue componenti funzionali espresse da fasi e cicli, il sonno appare pertanto un processo dipendente dall’attività di numerose strutture, differenti neurotrasmettitori e numerose neuromolecole, processo che ha differenti gerarchie (più essenziali sembrano le strutture filogeneticamente più antiche), ma anche con strette interdipendenze funzionali. Per questo motivo appare assai valida la saggia osservazione di Mancia che
sottolinea come la fisiologia del sonno sempre più si avvicina a quella dei grandi sistemi che sovrintendono i comportamenti più complessi, comprendenti, oltre le funzioni emotivo-affettive, anche quelle percettive e cognitive, chiaramente chiamate in causa dall’attività onirica che ricorre abbondante durante il sonno REM.
Leggi anche:
- Igiene del sonno: come aumentarla per dormire bene ed evitare l’insonnia
- Quante ore per notte devi dormire per essere riposato e stare bene?
- Per quanto tempo si può rimanere svegli senza impazzire o morire?
- Cos’è è il sonno, a che serve e perché è così importante dormire?
- Erezioni notturne involontarie, frequenti e continue: cosa indicano?
- Terrore notturno in adulti e bambini: cause psicologiche, epilessia, cure
- Bruxismo: sintomi, bite, prevenzione e cura
- Ipersonnie: tipi, classificazione, cause, sintomi, diagnosi, terapie
- Paralisi del sonno e allucinazioni ipnagogiche: cause, pericoli, rimedi
- Enuresi notturna in bambini, adolescenti e adulti: cause e terapia
- Morte in culla (SIDS): prevenzione, cause, sintomi e percentuale dei casi
- Perché si russa e quali sono i rimedi per smettere di russare? I pericoli dell’apnea ostruttiva del sonno
- Erezioni notturne dolorose, bisogno di urinare e ipertrofia prostatica
- Le 5 erezioni che ha l’uomo di notte: la tumescenza peniena notturna
- Erezioni del mattino: che indicano? Devono essere sempre presenti?
- Tumescenza clitorale notturna: l’erezione del clitoride durante la notte
- Apnea ostruttiva del sonno: cause, rischi, trattamenti e prevenzione
- Narcolessia: cause, sintomi, cure e terapia farmacologica
- Insonnia e apnee notturne: rischio di incidenti stradali triplicato
- Colpo di sonno improvviso alla guida: cause, conseguenze, rimedi
- Cataplessia: significato, cause e cura
- Nicturia: cos’è, cosa la provoca e come si cura
- Terapia non farmacologica dell’insonnia: dormire bene senza farmaci
- Farmaci sedativo-ipnotici usati per il trattamento di ansia ed insonnia
- Farmaci ipnotici per l’insonnia: quando usarli e quale scegliere
- Farmaci ipnotici: effetti indesiderati, rischi, sospensione, insonnia rebound
- Orologio biologico e disordini del ritmo circadiano sonno-veglia
- Ritmo circadiano: caratteristiche, durata, luce e melatonina
- Orologio biologico: come funziona e perché è importante
- Jet lag: cos’è, quanto dura, sintomi, rimedi e melatonina
- La morte per mancanza di sonno è davvero possibile? Per quanto tempo si può restare svegli?
- Disordini del ritmo circadiano: sindrome della fase di sonno ritardata
- Disordini del ritmo circadiano: sindrome della fase di sonno anticipata
- Disordini del ritmo circadiano: sindrome ipernictemerale
- Disordini del ritmo circadiano: sindrome del ritmo sonno-veglia irregolare
- Sindrome dei turnisti: l’orario di lavoro non rispetta il ritmo circadiano
- Jet lag e sindrome da salto di fusi orari: cause e terapie
- Mi alzo spesso di notte per urinare: quali sono le cause e le cure?
- Come cambia la temperatura corporea durante la giornata?
- Come cambia la temperatura corporea in base alla sede di misurazione?
- Come cambia la temperatura basale in base al ciclo mestruale?
- Sindrome da movimenti periodici delle gambe durante il sonno (PLMS)
- Sindrome delle gambe senza riposo e insonnia: sintomi e terapie
- Sindrome da sonno insufficiente: sintomi, diagnosi, evoluzione e terapie
- Insonnia psicofisiologica, sintomi, diagnosi e trattamento
- Insonnia da errata percezione del sonno (pseudoinsonnia): sintomi, diagnosi, cure
- Insonnia idiopatica: sintomi, diagnosi, conseguenze e cure
- Insonnia da allergia alimentare: sintomi, diagnosi e trattamento
- Insonnia da altitudine: cause, sintomi, diagnosi e terapia
- Insonnia da assunzione di cibo o bevande durante la notte nell’adulto
- Disturbo del sonno da adattamento: l’insonnia causata da eventi stressanti
- Sostanze ad azione stimolante sul sistema nervoso centrale che causano insonnia
- Sostanze che causano depressione del sistema nervoso centrale e insonnia
- Sostanze che causano insonnia: alcol, farmaci, metalli pesanti, inquinanti
- I diversi tipi di insonnia causati da patologie psichiatriche
- Insonnia da disturbi d’ansia, attacchi di panico, fobie, disturbo ossessivo compulsivo, stress post traumatico
- Insonnia da depressione (insonnia depressiva): sintomi e terapie
- Insonnia nel paziente con sindrome maniacale: sintomi e cure
- Insonnia nel paziente con schizofrenia: sintomi e trattamento
- Insonnia da inadeguata igiene del sonno nell’adulto: sintomi e cure
- Insonnia familiare fatale: diagnosi, trasmissione, cause, cure
- Disturbi dell’inizio del sonno “per associazione” nei bambini
- Insonnia e disturbo del sonno “limit setting” nei bambini
- Insonnia da assunzione di cibo o bevande durante la notte nel bambino
- Insonnia da inadeguata igiene del sonno in neonato, bambino e adolescente
- Psicoterapia, suggestione, ipnosi, meloterapia e training autogeno nell’insonnia
- Biofeedback nel trattamento non farmacologico dell’insonnia
- Cronoterapia nel trattamento non farmacologico dell’insonnia
- Fototerapia nel trattamento non farmacologico dell’insonnia
- Melatonina 1 e 2mg per insonnia: quando assumerla e controindicazioni
- Vi insegno le 12 regole d’oro per battere l’insonnia ed avere un sonno perfetto
- Valeriana in medicina: proprietà terapeutiche, fa male, controindicazioni
- E tu di che insonnia soffri? I cinque disturbi più frequenti sotto le coperte
- Come rilassarsi velocemente la sera subito prima di dormire in 20 passi
- Farmaci antidepressivi: cosa sono, a cosa servono e quali tipi esistono
- Sostanze naturali ad azione antidepressiva
- Disturbo ossessivo compulsivo: il trattamento con farmaci e terapia cognitivo-comportamentale è la migliore scelta
- Quali sono i benefici del sonno?
- Come interpretare correttamente i sogni
- Come imparare a controllare i sogni
- Come imparare a ricordare i sogni
- Le dieci cose che sicuramente non sai sui sogni
- Perché sogniamo? Perché ricordiamo alcuni sogni e altri no? Cos’è un sogno lucido e la fase REM?
- Che cos’è un “sogno lucido”?
- Sindrome delle gambe senza riposo: cause, sintomi, diagnosi, terapia
- Spasmi muscolari e mioclonie: cause, diagnosi e cura delle contrazioni involontarie
- Elettroencefalogramma: preparazione, alterazioni, costo, rischi
- Ipersonnia primaria e secondaria: cause, sintomi e rimedi
- Indossi sempre lo stesso pigiama? Ecco quali sono i rischi per la tua salute
- Mal d’auto, di mare, d’aereo: cinetosi, rimedi e farmaci per bambini ed adulti
- Dimmi come dormi e ti dirò chi sei
- Dormire senza struccarsi invecchia la pelle di dieci anni
- La donna deve dormire più dell’uomo: il cervello di lei lavora più di quello di lui
- Dormire poco fa ingrassare o dimagrire?
- Dormire col proprio cane fa riposare meglio
- Perché sbadigliamo e ci stiracchiamo? Perché lo sbadiglio è contagioso? Attrazione sessuale, noia ed altri misteri nascosti negli sbadigli
- Sono un sonnambulo: cause, sintomi, diagnosi e terapie
- Dormire dopo pranzo: la pennichella fa bene o male alla salute?
- Sognare topi, serpenti, gatti, sangue… Perché alcuni sogni sono ricorrenti e cosa significano?
- Troppo caldo per prendere sonno: i 14 trucchi per dormire bene in estate
- “Dottore, sono un sonnambulo del sesso!”
- Dormire col climatizzatore o il ventilatore fa male? Gli errori da non commettere
- Bruciore di stomaco: cosa mangiare, come dormire e rimedi naturali
- Sbadigliamo per sonno o per noia? Entrambe le risposte sono sbagliate
- Tumescenza peniena notturna, ovvero: le cinque erezioni che ha l’uomo di notte mentre dorme
- Mal di testa, sonnolenza, depressione, insonnia? E’ la sindrome di astinenza da caffeina
- La dieta per dormire bene, combattere l’insonnia e svegliarsi la mattina riposati
- Erezioni notturne dolorose, bisogno di urinare e ipertrofia prostatica
- Dormire poco ti consuma il cervello ma correre te lo ricostruisce
- Chi non riesce ad alzarsi presto la mattina è più intelligente e creativo
- Usare lo smartphone a letto prima di andare a dormire potrebbe ucciderti
- Differenze tra fase REM e NON REM del sonno
- Diventare più belli? basta dormirci su, ti spiego come!
- Stress: non basta dormire nel weekend per recuperare
- Beth, la “bella addormentata” esiste davvero
- Le cinque regole d’oro per dormire bene
- Bambini: meno dormono e più mangiano
- La morte per mancanza di sonno è davvero possibile? Per quanto tempo si può restare svegli?
- Ora solare, questa notte si cambia: a che ora e come spostare le lancette?
- Ora solare: cosa cambia nel nostro corpo (e nel nostro portafoglio)
- Male da ora solare: il drammatico primo lunedì dopo il cambio dell’orario
- Ora legale: quali sono gli effetti sul nostro corpo? Aumentano ictus e infarti, ma calano incidenti stradali
- Da ora solare a ora legale: come comportarsi coi bambini
- Nevrastenia (esaurimento nervoso): cause, diagnosi, cure
- Differenza tra astenia, ipostenia, miastenia, ipotonia, nevrastenia, iperstenia, ipertonia
- Stanchezza e debolezza: differenze, cause, rimedi e prevenzione
- Stanchezza pomeridiana tra lavoro e palestra: come ottenere il meglio dal vostro pomeriggio
- Sempre stanco e senza energia al lavoro? Ecco i rimedi
- Sei stanco e di cattivo umore: ecco i cibi che ti danno la carica ed i consigli per avere più energia
- Sei sempre stanco? Ecco le cause meno comuni che non immagineresti mai ed i rimedi
- Sonnolenza e stanchezza cronica: tutte le cause ed i rimedi
- Ansia da prestazione nello studio e nel lavoro: come superare le tue paure
- Lavori troppo? La tua salute è a rischio: ecco i 10 trucchi per faticare di meno a lavoro
- Combatti lo stress e ritrova il benessere psicofisico con il decalogo del buonumore
- Suicidarsi a causa del Minority Stress: quando appartenere ad una minoranza diventa fonte di discriminazione e sofferenza
- Differenze tra le varie scuole di psicoterapia: quale la più efficace?
- Scuola psicoanalitica (psicodinamica): l’efficacia della psicoanalisi
- Psicoterapia adleriana (o individualpsicologica)
- Psicoterapia cognitivo-comportamentale: lo schema comportamentale diventa sintomo
- Psicoterapia sistemico-relazionale: la famiglia converge sul paziente
- Psicoterapia psicosintetica: l’allontanamento dal Sé transpersonale
- Psicoterapia ericksoniana: l’ipnoterapia
- Psicoterapia funzionale: le alterazioni dei meccanismi psicofisiologici
- Depressione maggiore e minore, suicidio, diagnosi e cura: fai il test e scopri se sei a rischio
Lo Staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o unisciti al nostro gruppo Facebook o ancora seguici su Twitter, su Instagram o su Pinterest, grazie!
