 In risposta a stimoli lesivi, la prima linea di difesa dell’organismo è l’infiammazione. Le prime modificazioni che si verificano per generarla sono a carico dei vasi sanguigni di calibro minore cioè arteriole, venule e capillari. Si può dire che gli eventi che avvengono a livello vascolare sono volti a spingere la migrazione di cellule del sistema immunitario verso il distretto interessato dall’infezione o dalla lesione, a questo fenomeno si associa la perdita di proteine plasmatiche che fuoriescono dalla circolazione sanguigna invadendo i tessuti extravascolari.
In risposta a stimoli lesivi, la prima linea di difesa dell’organismo è l’infiammazione. Le prime modificazioni che si verificano per generarla sono a carico dei vasi sanguigni di calibro minore cioè arteriole, venule e capillari. Si può dire che gli eventi che avvengono a livello vascolare sono volti a spingere la migrazione di cellule del sistema immunitario verso il distretto interessato dall’infezione o dalla lesione, a questo fenomeno si associa la perdita di proteine plasmatiche che fuoriescono dalla circolazione sanguigna invadendo i tessuti extravascolari.
Vasocostrizione
La vasocostrizione con modificazioni di flusso è la prima manifestazione di un’infiammazione acuta. Generalmente è transitoria, incostante, dura da pochi secondi fino a qualche minuto e non è sempre presente. Interessa arteriole pre-capillari nelle immediate vicinanze della zona colpita e dipende dalla liberazione di sostanze vasocostrittrici di tipo catecolamminico (adrenalina, noradrenalina) e dalla serotonina.
Vasodilatazione o iperemia attiva
Inizia con il rilascio degli sfinteri arteriolari ed è successivamente sostenuta dall’apertura di nuovi letti capillari e dalla chiusura degli shunt arterovenosi fisiologicamente attivi a riposo. La vasodilatazione aumenta la quantità di sangue che rifornisce il tessuto leso (iperemia) ed è potenziata dall’aumento della permeabilità vascolare associata (vedi sotto). L’innesco ed il mantenimento della vasodilatazione sono dovuti al rilascio di mediatori rapidi come l’istamina o le prostaglandine e successivamente di mediatori lenti come IFN-γ, TNF-α, IL-1β, LPS, PAF. Questi ultimi stimolano l’espressione della sintetasi inducibile dell’ossido nitrico (iNos) a livello dell’endotelio vascolare. Al contrario l’istamina attraverso un aumento delle concentrazioni di calcio intracellulare determina l’attivazione (rapida) della NO sintasi costitutiva. Entrambe le isoforme di quest’ultimo enzima sono poi responsabili della sintesi di monossido di azoto (NO), un potentissimo vasodilatatore, che agisce sul muscolo liscio vascolare provocandone il rilassamento attraverso la mediazione di una guanilato ciclasi NO-dipendente. Quest’ultima genera guanosina monofosfato ciclica (cGMP), che a sua volta regola la funzione di numerosi enzimi e componenti cellulari coinvolti nella contrazione del muscolo liscio vascolare:
- inibisce la chinasi della catena leggera della miosina (MLCK)
- inibisce i recettori rianodici (RYR)
- attiva la proteina chinasi G (PKG), che a sua volta provoca un’attivazione della fosfatasi della catena leggera della miosina (MLCP: enzima antagonista di MLCK) e diminuisce le concentrazioni di calcio intracellulare (fondamentale per la contrazione del muscolo liscio)
- attiva la PGH-sintasi 1 e 2 favorendo il mantenimento dello stato infiammatorio e in ultima analisi della vasodilatazione.
L’aumento del calibro vascolare provoca:
- riduzione della velocità del flusso sanguigno (stasi): questa condizione favorisce il rolling leucocitario, rallenta l’eventuale diffusione di patogeni e favorisce l’adesione del complemento ai patogeni
- aumento della pressione transmurale con conseguente aumento della trasudazione vascolare e quindi del drenaggio linfatico.
L’aumento del drenaggio linfatico favorisce la presentazione antigenica.
Leggi anche:
- Shock settico e sepsi: sintomi, terapia, conseguenze, si può guarire
- Differenza tra sepsi e Sindrome da risposta infiammatoria sistemica (SIRS)
- Meningite: contagio, sintomi, vaccino, gravità e profilassi
- Endocardite: cause, sintomi, diagnosi e terapie
- Differenza tra batteri Gram negativi e Gram positivi
- Differenza tra batteri bacilli, cocchi, streptococchi e spirilli
Fase di rallentamento del circolo o iperemia passiva
Dopo la fuoriuscita di liquidi (essudazione) e di cellule (diapedesi) aumenta la viscosità del sangue, mentre diminuisce la velocità del flusso (stasi): nella zona infiammata i letti vascolari accolgono (come nelle fasi iniziali) più sangue della norma (iperemia), ma ciò è dovuto al rallentamento del circolo e al diminuito ritorno venoso (iperemia passiva) e non all’apertura di nuovi letti capillari (iperemia attiva). Le cause del diminuito ritorno venoso necessitano della compressione esercitata sulle venule da parte dell’edema, che ostacola così il passaggio di sangue provocando una riduzione della sua parte liquida ed un aumento relativo delle componenti solide.
Aumento della Permeabilità vascolare
Altro segno distintivo dell’Infiammazione acuta è l’aumento della Permeabilità vascolare a livello del microcircolo nell’area di lesione, dando luogo alla fuoriuscita di essudato, attraverso 5 meccanismi:
- Danno diretto da agente patogeno => endotelio distrutto e Membrana Basale (MB) lassa; è una risposta immediata e mantenuta a lungo in tutti i tipi di vasi, fino a riparazione del danno.
- Tipo Istamina => contrazione delle cellule endoteliali + disgiunzione delle unioni intercellulari; è immediata e transitoria per azione degli stessi mediatori che favoriscono la VD arteriolare, oltre a sostanze propriamente patogene, come l’endotossina, che stimola i fattori complemento (es: C5a), che a loro volta stimolano l’aumento della permeabilità, rinforzando la risposta flogistica.
- Lesione da Leucociti (o “Capillare tardiva”) => per extravasazione leucocitaria, durante la transepitelizzazione dei leucociti, e la secrezione di ulteriori mediatori cellulari e proteolitici; la risposta è tardiva e prolungata.
Inoltre, essendo principalmente a carico della componente capillare, il protrarsi del processo infiammatorio altera il normale equilibrio di scambi gassosi e idrici tra interstizio e microcircolo tale da produrre, insieme ad elementi di cui sopra e altri, la classica istolesività associata alla flogosi.
- Transcitosi (o citopempsi) aumentata => aumento della mobilità delle caveole transmembranarie, via preferenziale del plasma, favorendo il passo di materiale idrosolubile attraverso la MB.
- Neoangiogenesi => formazione di nuovi abbozzi vascolari durante la riparazione tissutale, delicati e facilmente emorragici fino alla formazione di nuove unioni intercellulari.
L’essudato è una miscela di liquidi plasmatici ricchi di proteine plasmatiche e/o cellule (per lo più infiammatori, come neutrofili e mononucleati, ma anche eritrociti), fuoriuscita dai vasi, per l’aumento della permeabilità capillare e iperemia attiva, ai tessuti o alle cavità sierose. Questo provoca edema infiammatorio. L’aumento della permeabilità è contemporaneo alla vasodilatazione che porta ad un aumento della massa di sangue presente e la pressione che esso esercita sulle pareti con aumento della permeabilità. L’aumento della pressione idrostatica e aumento della pressione colloido-osmotica determinano:
- dilatazione delle arteriole (aumento del flusso di liquidi verso l’esterno),
- apertura del letto capillare,
- dilatazione venulare che porta ad un aumento del flusso di liquidi verso l’esterno.
Leggi anche:
- Triade di Virchow: i tre fattori di rischio per la trombosi
- Chetosi: cos’è, da cosa è causata, sintomi e terapia in adulti e bambini
- Coagulazione intravascolare disseminata: cause e trattamenti
- Sepsi: cause, sintomi, diagnosi e terapie
- Differenza tra sepsi e setticemia
- Sindrome da disfunzione multiorgano: cause, sintomi, stadi e cure
- Coprocoltura feci per salmonella: perché e come si fa
- Batteriemia: cura, segni, sintomi, diagnosi ed antibiotici
Migrazione dei leucociti nella sede d’infiammazione
Le due categorie più importanti di leucociti coinvolte nel processo infiammatorio sono i granulociti neutrofili e i macrofagi (monociti nel flusso sanguigno), tuttavia, a seconda dello stimolo infiammatorio possono essere coinvolte altre classi di leucociti come i linfociti, i granulociti eosinofili e basofili o mastociti. Una funzione fondamentale dell’infiammazione è quella di permettere ai leucociti di giungere nella sede del danno e, quindi, di fagocitare agenti lesivi, uccidere batteri, degradare il tessuto necrotico e gli agenti estranei. Questo processo richiede l’attraversamento dell’endotelio vascolare delle venule post-capillari, che avviene in cinque fasi: la marginazione, il rotolamento, l’adesione, la diapedesi e la chemiotassi.
Normalmente il flusso sanguigno all’interno delle venule può essere descritto in modo approssimativo come composto di una colonna centrale in cui decorrono gli eritrociti e di uno spazio circostante prossimo alla parete endoteliale in cui decorrono prevalentemente leucociti di ogni tipo. La stasi ematica, una delle alterazioni che si verificano nel processo infiammatorio in conseguenza dell’aumento del calibro delle venule e dei letti capillari, favorisce la diminuzione di tensione delle pareti dei vasi sanguigni, ne consegue che molti più leucociti vengono confinati in prossimità dell’endotelio determinando quel fenomeno noto come marginazione. La stretta vicinanza con la parete endoteliale determina per i leucociti maggiori probabilità di adesione alla parete dell’endotelio qualora le cellule endoteliali e i leucociti stessi siano attivati da citochine. In tal caso i leucociti tendono a legarsi più volte ma in modo transitorio alle cellule endoteliali con legami deboli che vengono generalmente distrutti dal flusso sanguigno impedendo al leucocita l’adesione e facendolo rotolare lungo la parete del vaso (rotolamento). Il rotolamento dei leucociti è determinato da una classe di proteine di adesione note come selectine e dai loro ligandi specifici e complementari, espressi sia dall’endotelio che dai leucociti.
I leucociti esprimono L-selectina, i cui ligandi sulla superficie della membrana plasmatica delle cellule endoteliali sono GlyCam-1 (Glycosylation-dependent Cell adhesion molecule-1) e CD34. Le cellule endoteliali esprimono E-selectina e P-selectina, il cui ligando espresso dai leucociti è una proteina modificata con sialil-Lewis X. La P-selectina è conservata all’interno delle cellule endoteliali nei granuli chiamati corpi di Weibel-Palade e viene traslocata sul polo vasale membrana plasmatica sotto stimolazione da parte di istamina, trombina e fattore di attivazione delle piastrine (PAF). L’espressione delle altre selectine è mediata dalle citochine TNF (tumour necrosis factor) e IL-1, che vengono secrete prevalentemente dai macrofagi durante l’infiammazione. Il rotolamento dei leucociti li rallenta e permette la fase successiva, l’adesione, mediata da proteine eterodimeriche presenti sulla superficie leucocitaria note come integrine e dai loro ligandi sull’endotelio. Le integrine collocate sulla membrana dei leucociti si trovano normalmente in uno stato a bassa affinità. L’aumento dell’affinità di queste proteine avviene ad opera delle chemochine secrete anch’esse dai macrofagi e da altri leucociti durante il processo flogistico, che si legano ai proteoglicani dell’endotelio e quindi attivano le integrine dei leucociti. Le integrine più rappresentate sono LFA-1 e Mac-1 nei granulociti neutrofili, nei linfociti e nei monociti e VLA-4 nei granulociti eosinofili, monociti e linfociti. I loro ligandi sono rispettivamente ICAM-1 (inter-cellular adesion molecule-1) e VCAM-1 (vascular cell adhesion molecule-1), la cui espressione è stimolata ancora una volta da TNF e IL-1.
L’interazione tra integrine e i loro ligandi permette l’adesione dei leucociti alla parete endoteliale. Una volta adesi all’endotelio i leucociti sono attirati verso la sede d’infezione dal gradiente di concentrazione delle chemochine, prodotte da altre cellule mediatrici della risposta infiammatoria. Prima di giungere in sede, però, devono migrare attraverso l’endotelio e la sua lamina basale tramite un processo che prende il nome di diapedesi e che si verifica prevalentemente nelle venule. I leucociti migrano attraverso l’endotelio a livello delle giunzioni intercellulari tra cellule endoteliali che durante il processo flogistico risultano generalmente allargate a causa della retrazione delle cellule endoteliali indotta da istamina e ossido d’azoto oppure per danno diretto dell’endotelio. La migrazione avviene tramite diverse molecole di adesione, tra cui una delle più note è PECAM-1, espressa sia sulle giunzioni intercellulari dell’endotelio che sui leucociti. La membrana basale viene attraversata mediante la secrezione di collagenasi, che la degrada. Giunti nello spazio extracellulare migrano verso la sede di lesione seguendo il gradiente di concentrazione delle chemochine (chemiotassi) e facendosi strada attraverso la matrice extracellulare mediante legami con i proteoglicani.
Le chemochine sono un gruppo eterogeneo di molecole esogene ed endogene che attirano i leucociti. Tra le esogene vi sono ad esempio il lipopolisaccaride (LPS) di molti batteri, peptidi contenenti l’amminoacido N-formilmetionina (presente solo nei batteri e nei mitocondri); tra le endogene alcune citochine (come IL-8), proteine del complemento, leucotriene B4. Queste molecole raggiungono il leucocita e si legano a specifici recettori sulla sua superficie, appartenenti generalmente ai GPCR (recettori accoppiati a proteine G) i quali attraverso diverse vie di segnalazione generano secondi messaggeri che convergono sulla proteina Rho e simili GTPasi (Rac, cdc42). Queste proteine G sono fondamentali per la polimerizzazione dell’actina G nei filamenti di actina F e siccome questa proteina è la principale responsabile assieme alla miosina del movimento cellulare dei leucociti, ne permettono la migrazione sino alla sede di lesione. Generalmente l’actina costituisce l’impalcatura dei filopodi e ne permette l’estensione nel senso di marcia del leucocita. I filopodi permettono la trazione anteriore, mentre la miosina si dispone posteriormente aiutando a trascinare il grosso della cellula in avanti.
I leucociti (neutrofili, basofili, eosinofili, monociti) seguono tutti le stesse modalità di migrazione; i neutrofili, però, avendo vita più breve, vanno incontro ad apoptosi o NETosi e scompaiono in 24-48h. I monociti si trasformano in macrofagi e hanno vita più lunga. Eccezioni: infezioni da pseudomonas, infezionivirali, reazioni allergiche.
I leucociti hanno diverse funzioni:
- fagocitosi
- riconoscimento e adesione: i microrganismi sono riconosciuti se rivestiti da fattori (opsonine) che si legano a specifici recettori leucocitari (aumenta molto l’efficienza della fagocitosi);
- ingestione: dalla cellula si formano estensioni citoplasmatiche (pseudopodi) che avvolgono l’agente estraneo (fagosoma) e successivamente si forma il fagolisosoma
- uccisione: 02 dipendente e 02 indipendente;
- degradazione: dopo l’uccisione le idrolasi acide dei granuli dei leucociti degradano il batterio, il pH scende a valori di 4, ottimale per l’attivazione di questi enzimi.
- produzione di metaboliti dell’acido arachidonico;
- degranulazione e secrezione di enzimi lisosomiali;
- produzione di specie reattive dell’ossigeno (ROS);
- modulazione di molecole di adesione leucocitaria.
Leggi anche:
- Differenza tra virus e batteri: chi è più pericoloso? Diagnosi, sintomi e terapia
- I cinque segni cardinali dell’infiammazione
- Differenza tra infezione ed infiammazione: sono la stessa cosa?
- Differenza tra infezione acuta e cronica
- Morte cellulare: differenza tra necrosi, apoptosi ed autofagia
- Infestazione: cos’è, da cosa è causata, come si cura
- Differenza tra infezione ed infestazione
- Differenza tra infestazione interna ed esterna
- Differenza tra infiammazione cronica granulomatosa e non granulomatosa
- Differenza tra granulomi asettici (da corpo estraneo) e settici
- Linfonodi: cosa sono, come riconoscerli, quando sono pericolosi
- Linfonodo sentinella: cos’è e perché è importante in caso di cancro
- Biopsia del linfonodo sentinella: a che serve, perché è importante
- Sistema linfatico e linfonodi: anatomia e funzioni in sintesi
- Differenza tra cisti e linfonodo
- Differenza tra cisti, pseudocisti, ascesso ed empiema
- Differenza infiammabile, combustibile, comburente e facilmente infiammabile
- Differenza tra acidosi ed alcalosi, metabolica e respiratoria
- Peritonite: tipi, cause, sintomi, diagnosi e terapie
- Appendicite acuta e cronica: cause, sintomi e terapie
- Differenza tra empiema ed ascesso
- Virus e virioni: cosa sono, come sono fatti, come funzionano e come si riproducono
- HIV: dopo quanto si manifestano i sintomi? I 4 stadi dell’infezione
- HIV: sintomi iniziali in donne e uomini
- Differenza tra HIV e AIDS: sono uguali?
- Si muore di AIDS? Qual è l’aspettativa di vita?
- Sistema immunitario, immunità innata e specifica: riassunto, schema e spiegazione
- Immunodeficienza primaria e secondaria: sintomi, cause e terapie
- Immunità innata (aspecifica): barriere, infiammazione e complemento
- Immunità innata (aspecifica): neutrofili, macrofagi e linfociti natural killer
- Immunità specifica (acquisita): linfociti, T killer, T helper, T γδ, B ed anticorpi
- Immunità specifica (acquisita): memoria passiva, attiva ed immunizzazione
- Immunità specifica (acquisita) umorale e cellulare
- Differenza tra aterosclerosi e arteriosclerosi
- Differenza tra trombo, embolo, coagulo, embolia e trombosi
- Differenza tra arterie, vene, capillari, arteriole e venule
- Differenza tra arteriola afferente ed efferente: struttura e funzioni
- Differenza tra edema infiammatorio, non infiammatorio, essudato, trasudato, idropisìa e idrope
- Cos’è l’Idropisìa?
- Idrope: cause, tipi e terapia
- Cos’è l’edema, come e perché si forma?
- Differenza tra edema localizzato, generalizzato e sistemico
- Differenza tra insufficienza renale acuta, cronica e dialisi
- Crisi respiratoria acuta e rischio di morte: cosa fare?
- Apparato respiratorio: anatomia in sintesi, struttura e funzioni
- Asma bronchiale: spirometria e diagnosi differenzialeAsma bronchiale: spirometria e diagnosi differenziale
- Esofago e trachea: zona, anatomia, rapporti e differenze
- Spirometria diretta ed indiretta: come si esegue ed a cosa serve
- Differenza tra BPCO ed asma: terapia e sintomi comuni e diversi
- Broncopneumopatia cronica ostruttiva (BPCO): sintomi, diagnosi e cura
- Asma bronchiale in bambini e adulti: cause, sintomi e cura
- Differenza tra inspirazione e espirazione: l’atto respiratorio
- Tumore al polmone in chi non fuma: da cosa viene causato?
- Differenza tra ventilazione polmonare e alveolare: spazio morto anatomico e fisiologico
- Broncoscopia polmonare con biopsia: a cosa serve, fa male, è pericolosa?
- Tutti gli articoli sullo smettere di fumare
- Enfisema polmonare: sintomi, tipi, cause, diagnosi e terapia
- Alveoli polmonari: cosa sono e che funzioni svolgono?
- Enfisema polmonare: cure, complicazioni, quando chiamare il medico
- Spirometria diretta ed indiretta: come si esegue ed a cosa serve
- Che significa malattia autoimmune? Spiegazione ed esempi
- Parametri della spirometria: capacità, volumi, rapporti e flussi
- Polmoni: anatomia e funzioni in sintesi
- Differenza tra laringe, faringe e trachea
- Pneumotorace spontaneo primario, secondario ed iperteso: cause, sintomi, terapie
- Sangue dal naso (epistassi) in bambini e adulti: cause, rimedi naturali, cosa fare e cosa NON fare
- Differenza tra dispnea, apnea e tachipnea
- Differenza apnea statica, dinamica e profonda
- Differenza tra ipossiemia, ipossia ed anossia
- Differenza tra ipossiemia e ipercapnia
- Differenza tra insufficienza respiratoria di tipo 1 e 2
- Differenza tra costola incrinata e rotta
- Cos’è una costa? Differenza tra costole e coste
- Gabbia toracica: dove si trova, a che serve e da cosa è composta
- Costola rotta (frattura costale): sintomi, diagnosi e terapia
- Costola incrinata: sintomi, terapia e tempi di recupero
- Differenza tra costole e vertebre
- Muscoli respiratori volontari ed involontari
- Frattura costale multipla, volet costale e pneumotorace
Lo staff di Medicina OnLine
Se ti è piaciuto questo articolo e vuoi essere aggiornato sui nostri nuovi post, metti like alla nostra pagina Facebook o seguici su Twitter, su Instagram o su Pinterest, grazie!
 Con “anuria” si intende la sospensione quasi totale della produzione di urina, con diuresi inferiore a 100 ml nelle 24 ore. Tale sintomo è l’espressione di una brusca grave interruzione della funzione renale.
Con “anuria” si intende la sospensione quasi totale della produzione di urina, con diuresi inferiore a 100 ml nelle 24 ore. Tale sintomo è l’espressione di una brusca grave interruzione della funzione renale.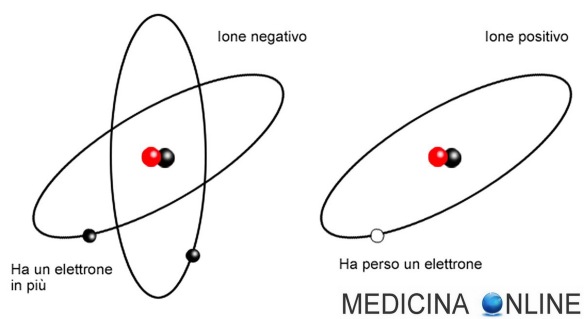 Secondo la teoria particellare della materia, l’atomo è la parte più piccola e indivisibile della materia. Tutti gli atomi presentano un numero atomico che corrisponde alla loro posizione nella tavola periodica. Il numero atomico esprime il numero di protoni presenti nel nucleo, ovvero il numero di particelle elementari che hanno carica elettrica positiva. In un atomo vi sono altrettanti elettroni con carica negativa. Pertanto gli atomi sono particelle elettricamente neutre, poiché lo stesso numero di cariche positive e negative si neutralizza a vicenda. Tuttavia se l’atomo cede o acquista elettroni dall’esterno perde la sua neutralità elettrica. Si ottiene uno ione positivo da un atomo che cede elettroni negativi. Se l’atomo acquista elettroni diventa uno ione con carica elettrica negativa.
Secondo la teoria particellare della materia, l’atomo è la parte più piccola e indivisibile della materia. Tutti gli atomi presentano un numero atomico che corrisponde alla loro posizione nella tavola periodica. Il numero atomico esprime il numero di protoni presenti nel nucleo, ovvero il numero di particelle elementari che hanno carica elettrica positiva. In un atomo vi sono altrettanti elettroni con carica negativa. Pertanto gli atomi sono particelle elettricamente neutre, poiché lo stesso numero di cariche positive e negative si neutralizza a vicenda. Tuttavia se l’atomo cede o acquista elettroni dall’esterno perde la sua neutralità elettrica. Si ottiene uno ione positivo da un atomo che cede elettroni negativi. Se l’atomo acquista elettroni diventa uno ione con carica elettrica negativa. Il termine sepsi indica una malattia sistemica causata dalla risposta dell’organismo all’invasione di tessuti, fluidi o cavità corporee normalmente sterili da parte di microrganismi patogeni o potenzialmente patogeni. Le complesse interazioni tra il microrganismo infettante, il sistema immunitario dell’ospite, le risposte infiammatorie e la coagulazione influenzano l’esito nella sepsi.
Il termine sepsi indica una malattia sistemica causata dalla risposta dell’organismo all’invasione di tessuti, fluidi o cavità corporee normalmente sterili da parte di microrganismi patogeni o potenzialmente patogeni. Le complesse interazioni tra il microrganismo infettante, il sistema immunitario dell’ospite, le risposte infiammatorie e la coagulazione influenzano l’esito nella sepsi. La coagulazione intravascolare disseminata (CID) o coagulazione vascolare disseminata è una gravissima sindrome clinica caratterizzata dalla presenza disseminata di numerosi trombi.
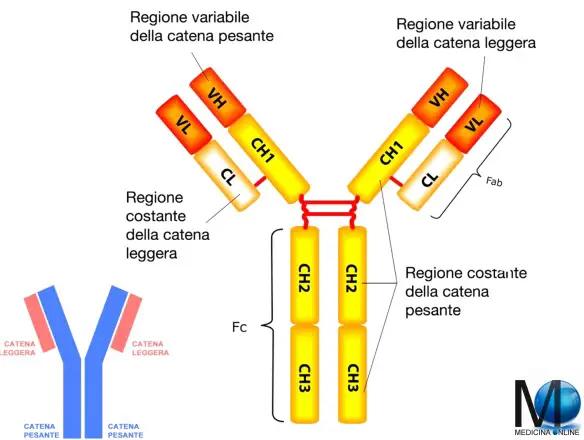
La coagulazione intravascolare disseminata (CID) o coagulazione vascolare disseminata è una gravissima sindrome clinica caratterizzata dalla presenza disseminata di numerosi trombi.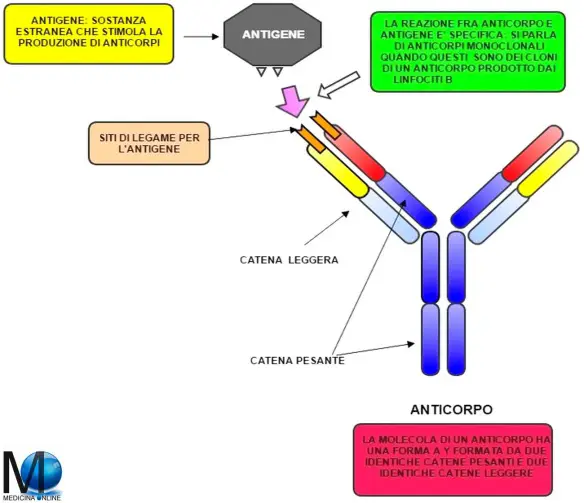 Gli antigeni sono sostanze in grado di essere riconosciute dal nostro sistema immunitario e possono essere classificati a seconda della loro classe:
Gli antigeni sono sostanze in grado di essere riconosciute dal nostro sistema immunitario e possono essere classificati a seconda della loro classe: